Membránový potenciálny rozdiel. Hlavné vlastnosti nervovej bunky. Bunková membrána ako elektrický reťazec
Článok pre hospodársku súťaž "BIO / MOL / TEXT": Potenciál ľudí je dôležitým fenoménom v živote všetkých buniek organizmu, a je dôležité vedieť, ako je vytvorený. Toto je však komplexný dynamický proces, ktorý je ťažké vnímať úplne, najmä pre študentov juniorských kurzov (biologické, lekárske a psychologické špeciality) a nepripravených čitateľov. Pri posudzovaní bodov je však celkom možné pochopiť jej základné detaily a etapy. Práca zaviedla pojem odpočinku potenciálu a zdôrazňuje hlavné etapy jeho formácie s použitím obrazových metaforov, ktoré pomáhajú pochopiť a zapamätať si molekulárne mechanizmy na tvorbu potenciálu odpočinku.
Membránové transportné konštrukcie - čerpadlá sodík-draslík - vytvárať predpoklady pre výskyt odpočinku. Predpokladom sú rozdielom v koncentrácii iónov na vnútorných a vonkajších stranách bunkovej membrány. Samostatne ukazuje rozdiel v koncentrácii sodík a rozdiel v koncentrácii podľa KALIA. Pokus o ióny draslíka (K +) zosúladiť ich koncentráciu na oboch stranách membrány vedie k jeho úniku z bunky a straty s nimi pozitívne elektrické poplatkyKvôli tomu je významne zvýšený všeobecný záporný náboj vnútorného povrchu buniek. Toto "draslíka" negatívnosť je väčšina potenciálu zvyšného potenciálu (-60 mV priemeru) a menšou časťou IT (-10 mV) je "výmena" negativity spôsobenej elektrinou samotnej iónovej výmennej čerpadla.
Zaoberáme sa viac.
Prečo potrebujeme vedieť, aký druh odpočinku a ako sa vyskytne?
Viete, čo je "elektrická energia zvierat"? Kde v tele vziať "biotoki"? ako Živé bunky.Nachádza sa vo vodnom prostredí, sa môže zmeniť na "elektrickú batériu" a prečo to okamžite vypúšťa?
Tieto otázky môžu byť zodpovedané len vtedy, ak viete, ako bunka vytvorí rozdiel v elektrických potenciáli (odpočinok) na membráne.
Je jasné, že na pochopenie toho, ako funguje nervový systém, je potrebné zistiť najprv, pretože jeho samostatná nervová bunka - neurón funguje. Hlavnou vecou je, že neurón je založený na pohybe elektrických nábojov prostredníctvom svojej membrány a vzhľadu elektrických potenciálov na membráne. Dá sa povedať, že Neuron, pripravuje sa na svoju nervovú prácu, najprv pokrýva energiu v elektrickej forme a potom ho používa v procese vodivosti a vysielania nervového vzrušenia.
Náš prvý krok k štúdiu práce nervového systému je teda pochopiť, ako sa elektrický potenciál objaví na membráne nervových buniek. Budeme sa zaoberať týmto spôsobom. tvorba potenciálu odpočinku.
Stanovenie koncepcie "potenciálu odpočinku"
Normálne, keď je nervová bunka vo fyziologickom pokoji a je pripravený na prácu, už mala redistribúciu elektrických obvinení medzi vnútornými a vonkajšími stranami membrány. Vďaka tomu vzniklo elektrické pole a na membráne sa objavil elektrický potenciál - membránový potenciál odpočinku.
Membrána sa teda ukáže, že bude polarizovaná. To znamená, že má iný elektrický potenciál vonkajších a vnútorných povrchov. Rozdiel medzi týmito potenciálom je možné zaregistrovať.
Toto môže byť overené, ak vstúpite do bunky Microelectrodu vo vnútri, pripojenú k registrujúcej inštalácii. Akonáhle elektróda spadá do bunky, okamžite získava nejaký trvalý elektronegatívny potenciál vzhľadom na elektródu umiestnenú v okolitej tekutinovej bunke. Veľkosť intracelulárneho elektrického potenciálu pri nervových bunkách a vláknach, napríklad obrie squid nervové vlákna, v pokoji, je asi -70 mV. Táto veľkosť sa nazýva membránový potenciál mieru (MPP). Vo všetkých bodoch axoplasmy je tento potenciál takmer rovnaký.
Nostdrachev a.d. a ďalšie. Začiatok fyziológie.
Viac fyziky. Makroskopické fyzické telá sú zvyčajne elektricky neutrálne, t.j. Sú v rovnakých súm obsiahnutých pozitívnymi aj negatívnymi poplatkami. Môžete si nabiť telo tým, že vytvorí nadbytok nabitých častíc jedného druhu v nej, napríklad trenie o inom telese, v ktorom je vytvorený prebytok opačných typov nábojov. Vzhľadom na prítomnosť elementárneho poplatku ( e.), plné elektrické nabitie akéhokoľvek tela môže byť reprezentované ako q. \u003d ± n × e.kde n je celé číslo.
Potenciálny odpočinok - Toto je rozdiel v elektrických potenciáli existujúcich na vnútorných a vonkajších stranách membrány, keď je bunka v stave fyziologického odpočinku. Jeho hodnota sa meria z vnútra bunky, je negatívny a je priemer -70 mV (Milvolt), aj keď v rôznych bunkách sa môže líšiť: od -35 mV na -90 mV.
Je dôležité vziať do úvahy nervový systém Elektrické poplatky nie sú znázornené elektrónmi, ako v bežných kovových drôtoch, ale iónmi - chemickými časticami, ktoré majú elektrický náboj. A všeobecne vo vodných roztokoch elektrický prúd Nie elektróny pohybujú, ale ióny. Preto všetky elektrické prúdy v bunkách a životné prostredie, ktoré ich obklopujú ión toki..
Takže z vnútra bunky sa nabíja negatívne a vonkajšia je pozitívna. Je to charakteristické pre všetky živé bunky, s výnimkou, s výnimkou červených krviniek, ktoré sú naopak, negatívne vonku vonku. Hovoriť konkrétnejšie, ukázalo sa, že kladné ióny (Na + a K + katióny) prevládajú okolo bunky a vo vnútri negatívnych iónov (organické kyslé anióny, ktoré nie sú schopné voľne pohybovať cez membránu ako NA + a K +) .
Teraz sme práve ponechali vysvetliť, ako to všetko presne ukázalo. Hoci, samozrejme, je to nepríjemné si uvedomiť, že všetky naše bunky okrem erytrocytov vyzerajú len pozitívne vonku, a vo vnútri sú negatívne.
Termín "negatívnosť", ktorý sa použijeme na charakterizáciu elektrického potenciálu vo vnútri bunky, bude užitočná pre nás pre jednoduchosť vysvetľovania zmien v úrovni odpočinku. V tomto termíne je cenné, že nasledujúce je intuitívne: čím viac negativity vo vnútri bunky - nižšie negatívny Potenciál sa posunie z nuly a menej negatívnosť - bližšie k negatívnemu potenciálu na nulu. To je oveľa jednoduchšie pochopiť, čo výraz "potenciál zvyšuje je presne to, čo znamená - zvýšenie absolútnej hodnoty (alebo" modul ") bude znamenať posun zvyšného potenciálu nadol z nuly a jednoducho" zvyšovanie "- potenciálneho posunu až do nuly. Termín "negatívnosť" nevytvára takéto problémy nejednoznačnosti porozumenia.
Podstatou tvorby potenciálu odpočinku
Poďme sa pokúsiť zistiť, kde je elektrický náboj nervových buniek odobratý, hoci nikto ich netrha, pretože fyzici robia vo svojich experimentoch s elektrickými poplatkami.
Tu výskumný pracovník a študent čaká jeden z logických pascí: vnútorná negativita bunky sa vyskytuje príchod z nepotrebných negatívnych častíc (anióny), ale naopak, kvôli straty niektorého počtu pozitívnych častíc (Katióny)!
Takže kde sú pozitívne nabité častice z klietky? Dovoľte mi pripomenúť, že opustil bunku a sodíkové ióny nahromadené mimo vonkajšej strany - Na + - a draslík - K +.
Hlavné tajomstvo vzhľadu negativity vo vnútri bunky
Túto tajomstvo okamžite otvoríme a hovoríme, že bunka je zbavená častí svojich pozitívnych častíc a je negatívne v dôsledku dvoch procesov:
- najprv si vymieňa "jeho" sodík "na" niekoho iného "draslíka (áno, jeden pozitívny ióny na strane druhej, rovnaké pozitívne);
- potom dôjde k úniku týchto "netrpezlivých" pozitívnych iónov draslíka, spolu s ktorými sú pozitívne návyky vysušené z bunky.
Tieto dva procesy, ktoré potrebujeme vysvetliť.
Prvá etapa vytvorenia vnútornej negativity: Exchange Na + na K +
V membráne nervovej bunky neustále pracuje proteín výmenní čerpadlá(adenosyntriphosfatáza, alebo Na + / K + -atfáza) vložená do membrány. Zmenia "vlastné" sodíkové bunky na vonkajšie "cudzinec" draslík.
Ale koniec koncov, pri výmene jedného kladného poplatku (NA +), neexistuje žiadny nedostatok pozitívnych poplatkov v bunke, aby sa vyskytla na inom kladnom nábore (K +)! Správny. Ale však kvôli tomu je v bunke veľmi málo sodíkových iónov, pretože takmer všetky vynechali. A súčasne je bunka ohromená iónmi draslíka, ktoré molekulárne čerpadlá boli prečerpané do nej. Ak by sme sa mohli pokúsiť ochutnať cytoplazmu bunky, všimli by sme, že v dôsledku prevádzky výmenných čerpadiel sa ukázalo z slanej v horkej slanej, pretože chlorid sodný tuhá chuť bola nahradená ťažkou chuťou koncentrovaného Roztok chloridu draselného. V bunke dosahuje koncentrácia draslíka 0,4 mol / l. Roztoky chloridu draselného v rozsahu 0,009-0,02 mol / l majú sladkú chuť, 0,03-0,04 - horký, 0,05-0,1 - horký a slaný a od 0,2 a vyššej - komplexná chuť pozostávajúca zo slanej, horkej a kyslá .
Dôležité je, že výmena sodíka pre draslík - nerovnavo. Pre každú bunku tri sodíkové ióny Dostane všetko dva ióny draslíka. To vedie k strate jedného kladného obvinenia s každou iónomeničnou činnosťou. Takže v tejto fáze v dôsledku nerovnakej výmeny bunky stráca viac "plusy", než prichádza na oplátku. V elektrických termínoch je to približne -10 mV negativity vo vnútri bunky. (Ale pamätajte, že stále potrebujeme vysvetlenie pre zostávajúce -60 mV!)
Aby bolo ľahšie zapamätať si prácu výmenných čerpadiel, je obrazne vyjadrená takto: "Bunka miluje draslík!"Preto bunka tiež línie draslíka k sebe, napriek tomu, že je to plné. A tak nevýhodne zdieľa na sodík, čím sa získa 3 ióny sodíka pre 2 ióny draslíka. A tak trávi na tejto výmennej energii ATP. A ako to trávi! Až 70% všetkých spotreby energie neurónov môže ísť do práce sodík-draslíkových čerpadiel. (To je to, čo robí láska, nechaj ju ešte nereá!)
Mimochodom, je zaujímavé, že bunka sa nenarodila s hotovým potenciálom odpočinku. Stále je potrebné vytvoriť. Napríklad pri diferenciácii a zlúčení myoblastov, potenciál ich membrány sa líši od -10 do -70 mV, t.j. Ich membrána sa stáva negatívnejšou - polarizuje v procese diferenciácie. A v experimentoch na multipotentných mezenchymálnych bunkách stromálnej kostnej drene, umelou depolarizáciou, proti odpočinku potenciálu a redukciu negatívnej bunky, dokonca inhibované (utláčané) bunkové diferenciácie.
Obrazovité rozprávanie, môže byť vyjadrené takto: vytvorenie potenciálu mieru, bunkovej "obvinenia s láskou." Toto je láska k dve veci:
- láska bunky do draslíka (preto je bunka násilne vypnutá k nemu);
- láska draslíka na slobodu (preto draslík opustí zachytenú bunku).
Už sme vysvetlili mechanizmus saturácie CAGE buniek (to je práca výmenných čerpadiel) a mechanizmus starostlivosti o draslík z bunky je vysvetlený nižšie, keď sa obrátime na opis druhého stupňa vytvorenia intracelulárnej negativity. Výsledok činností membránových iónových čerpadiel v prvej fáze tvorby potenciálu mieru je teda: \\ t
- Nedostatok sodíka (Na +) v bunke.
- Prebytok draslíka (K +) v bunke.
- Vzhľad na membráne slabého elektrického potenciálu (-10 mV).
Môže sa povedať: V prvej fáze, membránové iónové čerpadlá vytvárajú rozdiel koncentrácií iónov, alebo gradient (diferenciál) koncentrácie, medzi intracelulárnym a extracelulárnym médiom.
Druhá etapa vytvorenia negativity: Únik iónov K + z bunky
Takže, čo začína v klietke po membránovi čerpadlá s sodným-draslíkom pracujú s iónmi?
Kvôli deficitu sodíka vo vnútri bunky je tento ión ustrovaný na sebe. ponáhľať sa: Rozpustené látky vždy majú tendenciu zosúladiť ich koncentráciu v celom objeme roztoku. Ale tento sodík sa tesňuje zle, pretože iónové sodíkové kanály sú zvyčajne uzavreté a otvorené len za určitých podmienok: pod vplyvom špeciálnych látok (vysielačov) alebo so znížením negativity v bunke (depolarizácia membrány).
Bunka má zároveň prebytok iónov draslíka v porovnaní s vonkajším médiom - pretože membránové čerpadlá násilne čerpá do klietky. A tiež sa snaží vyrovnať svoju koncentráciu vo vnútri a vonku, usiluje sa naopak, ukončite klietku. A to sa ukáže!
Ióny draslíka K + Nechajte bunku pod pôsobením chemického gradientu ich koncentrácie rôzne strany Membrány (membrána je oveľa priepustná pre K + ako na NA +) a s nimi nesú pozitívne poplatky. Z tohto dôvodu sa negativita zvyšuje vo vnútri bunky.
Stále je dôležité pochopiť, čo sú sodíkové a draselné ióny "nevideli" navzájom, reagujú len "na seba". Tí. Sodík reaguje na koncentráciu sodíka, ale "nevenuje pozornosť" k tomu, koľko draslíka okolo. Naopak, draslík reaguje len na koncentráciu draslíka a "nevšimne si" sodík. Ukazuje sa, že pochopiť správanie iónov, je potrebné oddelene zvážiť koncentrácie sodíkových a draslíkových iónov. Tí. Je potrebné samostatne porovnať koncentráciu sodíka vo vnútri a mimo bunky a oddelene - koncentrácia podľa draslíka vo vnútri a mimo bunky, ale nemá zmysel porovnať sodík s draslíkom, ako sa to stane, vykonáva sa v učebniciach.
Podľa zákona o zosúlaďovaní chemických koncentrácií, ktoré pôsobia v roztokoch, sodík "chce" zvonku, aby vstúpil do bunky; Tam je tiež elektrická sila rovnakým spôsobom (ako si pamätáme, cytoplazmus je negatívne nabitá). More, ktorý chce, ale nemôže, pretože membrána v obvyklom štáte to chýba. K dispozícii sú sodíkové iónové kanály sú k dispozícii v membráne sú normálne. Ak ešte stále prichádza trochu, bunka si ho okamžite vymieňa na vonkajšie draslík pomocou jeho čerpadiel výmenných čerpadiel sodíka. Ukazuje sa, že sodíkové ióny prechádzajú klietkou, akoby tranzitu a nemajú v ňom oneskorenie. Preto je sodík v neurónoch vždy nedostatok.
Ale draslík len môže ľahko opustiť bunku vonku! Je plne v klietke a ona ju nemôže držať. Vychádza z špeciálnych kanálov v membráne - "kanály úniku draslíka", ktoré sú normálne otvorené a produkujú draslík.
K + únikové kanály sú neustále otvorené pri normálnych režimoch membrány zvislosti mieru a vykazujú výbušniny aktivity počas nožov membránového potenciálu, ktorý trvá niekoľko minút a sú pozorované so všetkými potenciálnymi hodnotami. Posilnenie úniku K +-topov vedie k hyperpolarizácii membrány, zatiaľ čo ich potlačenie je depolarizácia. ... Avšak existencia kanálového mechanizmu zodpovedného za úniku prúdy na dlhú dobu zostal. Len teraz sa ukázalo, že únik draslíka je prúd prostredníctvom špeciálnych draslíkových kanálov.
Zephyrov A.l. A SITDIKOVA G.F. Iónové kanály excitabilné bunky (štruktúra, funkcia, patológia).
Z chemických - na elektrické
A teraz - opäť najdôležitejšia vec. Musíme sa vedome pohybovať z pohybu chemické častice Pohyb elektrické poplatky.
Draslík (K +) je pozitívne nabitý, a preto, keď vyjde z klietky, sa z neho nevyžaduje nielen sám, ale aj kladný náboj. Za ním, z vnútra buniek na membránový úsek "mínusy" - negatívne poplatky. Ale nemôžu uniknúť cez membránu - na rozdiel od iónov draslíka - pretože Pre nich nie sú žiadne vhodné iónové kanály a membrána ich nenechajte ujsť. Zapamätajte si zostávajúce -60 mV negativity, ktoré zostáva -60. To je najviac časť membránového odpočinku, ktorý je vytvorený únikom draslíkových iónov z bunky! A to je väčšina z potenciálu odpočinku.
Pre túto časť súčasťou potenciálu zvyšku existuje aj špeciálny názov - potenciál koncentrácie. Koncentračný potenciál - Toto je súčasť potenciálu zvyšku, vytvorený nedostatkom pozitívnych poplatkov vo vnútri bunky, tvorený únikom z jej pozitívnych iónov draslíka.
No, teraz niektorí fyzici, chémia a matematika pre milovníkov presnosti.
Elektrické sily sú spojené s chemikáliami podľa Goldmanovej rovnice. Jeho špeciálny prípad je jednoduchšia nervová rovnica, ktorou vzorec, ktorý môže vypočítať transmembránový difúzny rozdiel potenciálov na základe rôznych koncentrácií iónov jedného druhu na rôznych stranách membrány. Takže, poznávať koncentráciu iónov draslíka mimo a vo vnútri bunky, môžete vypočítať rovnovážny potenciál draslíka E. K:

kde E.k - rovnovážny potenciál R. - konštanta plynu, \\ t T. - absolútna teplota, F. - Trvalá faraday, K + externé a K + vnútorné koncentrácie iónov do + vonku a vo vnútri bunky. Podľa vzorca je zrejmé, že koncentrácia iónov jedného druhu sa porovnáva s výpočtom potenciálu - K +.
Presnejšou konečnou hodnotou celkového difúzneho potenciálu, ktorý je vytvorený únikom niekoľkých typov iónov, sa vypočíta pomocou vzorca Goldman-Hodgkin-Katza. Zohľadňuje to, že potenciál starostlivosti závisí od troch faktorov: (1) polaritu elektrického náboja každého iónu; (2) permeabilita membrány Ročník pre každý ión; (3) [Koncentrácie zodpovedajúcich iónov] vnútri (vnútorné) a mimo membrány (externé). Pre diabrane Axon squid sám Ročník K: Pička :P. \\ t Cl \u003d 1: 0,04: 0,45.
Záver
Tak, pot zvyšku niak sa skladá z dvoch častí:
- -10 mVktoré sa získavajú z "asymetrickej" práce membránového výmenného kurzu výmenníka (po tom všetkom, čerpá pozitívne návyky z bunky (NA +), než stiahne späť s draslíkom).
- Druhou časťou je celoročne vysušené draselné bunky, s pozitívnymi nábojmi. Jeho príspevok - hlavné: -60 mV.. Súčet, to dáva požadovaný -70 mV.
Čo je zaujímavé, draslík prestane opustiť bunku (presnejšie, jeho vstup a výťažok je vyrovnaný) len na úrovni negatívnosti bunky -90 mV. V tomto prípade chemické a elektrické sily, ktoré tlačia draslík cez membránu, ale vedie ho do opačné strany. To však presahuje neustále unikajúce v bunkovej sodnej, ktorá prináša s ním pozitívnymi poplatkami a znižuje negatívnosť, pre ktoré boje draslíka. A v dôsledku toho bunka udržiava rovnovážny stav pri -70 mV.
Teraz je nakoniec vytvorený rekreačný potenciál odpočinku.
Schéma práce NA + / K + -atphase Vizuálne ilustruje "asymetrickú" výmenu NA + na K +: Čerpanie z nadmerného "plus" v každom cykle enzýmovej práce vedie k zápornému náboja vnútorného povrchu membrány. Čo sa nepovedalo v tomto videu, takže je to skutočnosť, že ATPAZ je zodpovedný za menej ako 20% potenciálu zvyšku (-10 mV): Zostávajúca "negatívnosť" (-60 mV) sa objavuje kvôli výstupu Bunka cez "Kaliya potápačské kanály" K +, ktoré sa snažia vyrovnať svoju koncentráciu vo vnútri bunky a mimo neho.
Literatúra
- Jacqueline Fischer-Lougheed, Jian-Hui Liu, Estelle Espinos, David Mordasini, Charles R. Bader, et. Al .. (2001). Ľudská myoblastová fúzia vyžaduje expresiu funkčného vnútorného usmerňovača KIR2.1 kanálov. J Cell Biol.. 153 , 677-686;
- Liu J.h., BiJlenga P., Fischer-Lougheed J. a kol. (1998). Úloha vnútorného usmerňovača K + prúdom a hyperpolarizácie v ľudskej fúzii myoblastov. J. Physiol. 510 , 467–476;
- Sarah Sundellacruz, Michael Levin, David L. Kaplan. (2008). Membránový potenciál kontroluje adipogénne a osteogénne diferenciácie mezenchymálnych kmeňových buniek. Plos jeden. 3 , E3737;
- Pavlovskaya m.v. a Mamyakin A.I. Elektrostatika. Dielektrika a vodiče v elektrickom poli. Trvalý prúd / Elektronický manuál Podľa všeobecného priebehu fyziky. Petrohrad: Štátna elektrotechnická univerzita v St. Petersburg;
- Nosdraachev A.D., Bazhenov Yu.I., Banannikova I.A., Bauev A.S. a iné. Spustiť fyziológiu: Učebnica pre univerzity / ed. Acad. Peklo. Nostrhachev. Petrohrad: LAN, 2001. - 1088 p.;
- Makarov A.M. a Luneva L.A. Základy elektromagnetizmu / fyziky v technická univerzita. T. 3;
- Zephyrov A.l. A SITDIKOVA G.F. Iónové kanály excitabilné bunky (štruktúra, funkcia, patológia). Kazaň: Art Cafe, 2010. - 271 p.;
- Vlasť t.g. Senzorická analýza potravinárskych výrobkov. Návod pre študentov univerzít. M.: Academy, 2004. - 208 p.;
- Colman ya. A REM K.-G. Vizuálna biochémia. M.: Mir, 2004. - 469 p.;
- Schulgovsky v.v. Základy neurofyziológie: tutoriál pre vysokoškolských študentov. M.: Aspekt Press, 2000. - 277 s ..
Každá živá bunka je pokrytá semipermeabilnou membránou, cez ktorú sa vykonáva pasívny pohyb a aktívna volebná preprava pozitívnych a negatívne nabitých iónov. V dôsledku tohto prenosu medzi vonkajším a vnútorným povrchom membrány je rozdiel v elektrických nábojoch (potenciály) - membránový potenciál. Existujú tri rôzne prejavy membránového potenciálu - membránový potenciálny mier, miestny potenciálalebo miestna odpoveď, I. akčný potenciál.
Ak nie sú na klietke žiadne vonkajšie stimuly, membránový potenciál je dlhá uložená konštanta. Membránový potenciál takejto pokojovej bunky sa nazýva membránový potenciál odpočinku. Pre vonkajší povrch bunkovej membrány je potenciál odpočinku vždy pozitívny a pre vnútorný povrch bunkovej membrány je vždy negatívny. Je zvyčajné merať potenciál odpočinku na vnútornom povrchu membrány, pretože Iónová kompozícia cytoplazmy buniek je stabilnejšia ako intercelulárna tekutina. Množstvo potenciálu odpočinku je relatívne konštantné pre každý typ bunky. Pre priečne svalové bunky sa pohybuje od -50 do -90 mV a pre nervové bunky od -50 do -80 mV.
Príčiny výskytu potenciálu odpočinku sú rôzne koncentrácie katiónov a aniónov vonku a vo vnútri bunky volebná priepustnosť Pre nich, bunková membrána. Cytoplazmus odpočítavania nervóznej a svalovej bunky obsahuje asi 30-50-krát viac draslíkových katiónov, 5-15-krát menej sodíkových katiónov a 10-50-krát menej chlórových aniónov ako extracelulárna kvapalina.
V pokoji, takmer všetky sodné bunky bunkovej membrány sú zatvorené a väčšina draslíkových kanálov je otvorená. Kedykoľvek sa na otvorený kanál beží ióny draslík, prechádzajú cez membránu. Keďže draselné iónové bunky sú oveľa väčšie, osmotická sila ich tlačí z bunky. Draselné katióny zvýšili pozitívny náboj na vonkajšom povrchu bunkovej membrány. V dôsledku výstupu iónov draslíka z bunky by ich koncentrácia vo vnútri a von z bunky čoskoro bola rovnaká. Tým sa zabráni odpudzovaniu elektrickej sily pozitívnych iónov draslíka z pozitívne nabitého vonkajšieho povrchu membrány.
Čím viac je hodnota pozitívneho náboja na vonkajšom povrchu membrány, tým ťažšie ísť z cytoplazmy cez membránu. Ióny draslíka opustia bunku, až kým sa elektrická responsion sila nechá rovná výkonu osmotického tlaku na +. S touto úrovňou potenciálu na membráne, vstup a výstup draslíkových iónov z bunky sú v rovnováhe, takže elektrický náboj na membráne je v tomto momente povolaný potenciál kalive Equilibrium. Pre neuróny sa rovná -80 až -90 mV.
Pretože takmer všetky sodíkové membrány sodíkové kanály sú uzavreté, potom Na + ióny prichádzajú do bunky v koncentračnom gradiente v menších množstvách. Znovu uhradia stratu pozitívneho náboja vnútorného média bunky spôsobené výstupom iónov draslíka, ale nemôže významne kompenzovať túto stratu. Preto prenikanie do bunky (úniku) sodíkových iónov vedie len na menšie zníženie membránového potenciálu, v dôsledku čoho je rekrebitný potenciál odpočinku v porovnaní s rovnovážnym potenciálom draslíka mierne menšiu hodnotu.
Teda katióny draslíkových katiónov spolu s nadbytkom katiónov sodíka v extracelulárnej tekutine vytvárajú pozitívny potenciál na vonkajšom povrchu membrány pokojovej bunky.
V stave zvyšku je plazmatická membránová bunka dobre priepustná pre chlórové anióny. Chlórové anióny, ktoré sú väčšie v extracelulárnej tekutine, difúzujú vo vnútri bunky a nesú negatívny náboj s nimi. Úplná vyrovnanie koncentrácií chlórových iónov vonku a vo vnútri bunky sa nevyskytuje, pretože To zabraňuje výkonu elektrického vzájomného odpudania rovnakých mien. Vytvorený potenciál chlóru V ktorom je vstup do chlórových iónov do bunky a ich výstup z nej je rovnováha.
Bunková membrána je prakticky nepriepustná pre veľké anióny organických kyselín. Preto zostávajú v cytoplazme a spolu s prichádzajúcim chlórovým aniónom poskytujú negatívny potenciál na vnútornom povrchu membrány pokojovej nervovej bunky.
Najdôležitejším významom membránového odpočinku je, že vytvára elektrické pole, ktoré ovplyvňuje makromolekuly membrány a dáva im na nabité skupiny určitú polohu v priestore. Je obzvlášť dôležité, aby toto elektrické pole určuje uzavretý stav aktivačnej brány sodíkových kanálov a otvoreného stavu ich inaktivačných brán (obr. 61, A). To zaisťuje stav klietky a jeho pripravenosť na vzrušenie. Dokonca aj relatívne malý pokles potenciálu potenciálu zvyšok otvorí aktiváciu "bránu" sodíkových kanálov, ktorá zobrazuje bunku zo stavu odpočinku a spôsobuje vzrušenie.

Na signál z predchádzajúcej bunky do nasledujúceho, neurón vytvára elektrické signály v sebe. Vaše pohyby s očami pri čítaní tohto odseku, pocit mäkkej stoličky pod korisť, vnímanie hudby zo slúchadiel a oveľa viac sú založené na skutočnosti, že stovky miliárd elektrických signálov prechádzajú do vás. Takýto signál sa môže narodiť v mieche a ísť na špičku prsta na dlhej axóne. Alebo môže prekonať zanedbateľnú vzdialenosť v hĺbke mozgu, obmedzená na limity interneyrónu s krátkymi procesmi. Akýkoľvek neurón, ktorý dostal signál, to prebieha cez jeho telo a vyrastá a tento signál má elektrickú povahu.
Späť v roku 1859, vedci boli schopní merať rýchlosť, s ktorou sa tieto elektrické signály prenášajú. Ukázalo sa, že elektrina prenášaná živým axonom je zásadne odlišná od elektrického prúdu v kovoch. Metalickým drôtom sa elektrický signál prenáša rýchlosťou v blízkosti rýchlosti svetla (300 000 kilometrov za sekundu), pretože v kovu je mnoho voľných elektrónov. Napriek tejto rýchlosti však signál relatívne oslabuje, prekonávajú dlhé vzdialenosti. Ak sa na Acesons, signály boli prenášané rovnakým spôsobom, ktorý sa prenášajú v kovoch, potom nervový impulz, ktorý pochádza z nervového konca v koži palca vašej nohy, by úplne v prdeli, bez toho, aby sa dosiahol váš mozog - elektrické Odolnosť organickej hmoty je príliš veľká a signál je príliš slabý.
Štúdie ukázali, že elektrická energia sa prenáša na axóny oveľa pomalšie ako na drôtoch, a že v srdci tohto prenosu leží neznámy mechanizmus skôr, v dôsledku čoho signál sa vzťahuje rýchlosť približne 30 metrov za sekundu. Elektrické signály, ktoré bežia okolo nervov, na rozdiel od signálov, ktoré sa dostanú na drôty, neoslabujú pozdĺž ich pohybu. Dôvodom je, že nervové zakončenia neprechádzajú samotným signálom pasívne, jednoducho umožňujú, aby sa nabité častice k dispozícii v nich, aby ich navzájom prenášali. Sú v každom z jeho bodu aktívny vysielač tohto signálu, ktorý ho prenáša, a detailný popis Tento mechanizmus si bude vyžadovať samostatnú kapitolu. Obetovanie vysokej rýchlosti nervových impulzov, vďaka aktívnemu prenosu signálu neurónov, dostáva záruku veľký prst Signál nôh sa dostane do miechy, nie je vôbec oslabená.

Dodržiavať priechod elektrickej excitačnej vlny alebo akčný potenciál (akčný potenciál ['ækʃʃn Pə'Tenʃʃl]), v živej klietke, pomerne jednoduché zariadenie: jeden koniec tenkého kovového drôtu je umiestnený na vonkajšom povrchu axónu dotykového neurónu kože a druhý sa dodáva na rekordér, kreslenie line, keď Signál je získaný a dole pri oslabení. Každý dotyk kože spôsobuje jeden alebo niekoľko možností pôsobenia. Ak dôjde k každému potenciálu, rekordér čerpá úzky dlhý vrchol.
Potenciál pôsobenia senzorického neurónu trvá len asi 0,001 sekúnd a obsahuje dve fázy: rýchle zvýšenie, dosiahnutie vrcholu a potom takmer ako rýchly rozpad extácie, ktorý vedie k pôvodnej polohe. A potom rekordér informuje neočakávanú skutočnosť: všetky potenciály činností vznikajúcich v rovnakej nervovej bunke sú približne rovnaké. Je možné vidieť na obrázku vľavo: Všetky vrcholy nakreslené rekordérom majú približne rovnaký tvar a amplitúda, bez ohľadu na to, aké silné alebo dlhé bolo dotyk kože, čo ich spôsobilo. Slabé hladiace alebo hmatateľné zástrčky budú prevedené potenciálom pôsobenia tej istej hodnoty. Akčný potenciál je trvalý signál, s výhradou zásady "All alebo Nothing": Po prekročení dráždivého určitej prahovej hodnoty sa vždy vyskytuje približne o rovnakom signáli, nie viac a nie menej ako obvykle. A ak je stimul menší ako prahová hodnota, signál nebude vždy prenášaný: napríklad môžete tak ľahko dotknúť pokožky pera špičkou, že tento dotyk nebude cítiť.
Princíp "všetko alebo nič" v vzniku akčného potenciálu spôsobuje nové otázky. Ako sa NEUND NEURON uvádza silu stimulu - silný alebo slabý tlak, svetlé alebo slabé svetlo? Ako uvádza trvanie dráždivého? Nakoniec, ako sa neuróny vyznačujú jedným typom zmyslových informácií z druhej - ako sa rozlišujú dotyk od bolesti, svetla, vône alebo zvuku? A ako rozlišujú zmyslové informácie na vnímanie z informácií o motoroch pre akciu?

Evolúcia vyriešila otázku, ako nahlásiť silnú stimulu, s použitím použitia rovnakého typu signálov rovnakej hodnoty: táto sila sa stanoví frekvencia (Frekvencia ['friːkwənsɪ]), s ktorými sú emitované potenciály účinku. Slabý stimul, napríklad mierny dotyk smerom k ruke, vedie k emisii len dvoch alebo troch potenciálov pôsobenia za sekundu, zatiaľ čo silný tlak, ako keď je štrbina alebo šok, môže spôsobiť frontu stoviek potenciálov za sekundu . V tomto prípade sa trvanie pocitu určuje trvanie výskytu potenciálov pôsobenia.
Sú neuróny rôznych elektrických kódov, rozprávajú mozog, ktorý nesú informácie o rôznych stimuloch, ako je bolesť, svetlo alebo zvuk? Ukázalo sa, že nie je! To je prekvapujúce, ale medzi potenciálom akcie generovanej neurónmi z rôznych senzorických systémov (napríklad vizuálne alebo hmatové), rozdiel je veľmi zanedbateľný! Príroda a povaha potencie teda nezávisí od rozdielov v potenciáloch akcie (ktorá otvára skôr vzrušujúcu perspektívu na premýšľanie o téme "Matrix" z filmu rovnakého mena). Neurón, vysielanie sluchových informácií, je usporiadané rovnakým spôsobom ako neurón z vizuálneho nervového reťazca, a oni sú strávené rovnakým akčným potenciálom, rovnakým spôsobom. Bez vedomia, niektorí nervový reťazec vlastní špecifický neurón, nie je možné určiť, ktoré informácie nesie.
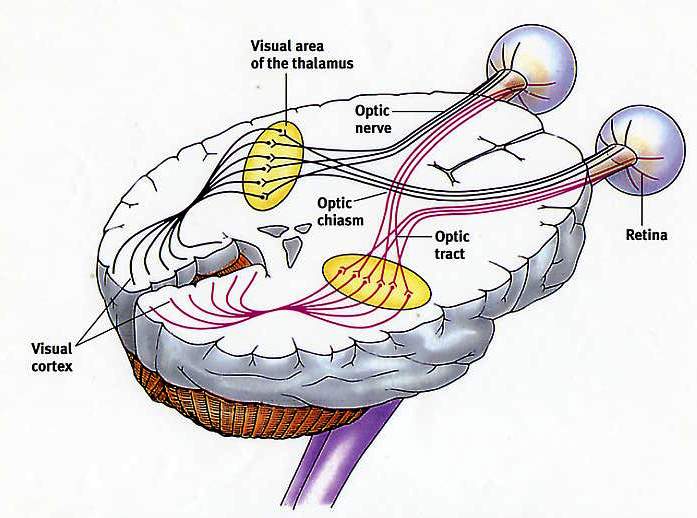
Povaha prenášaných informácií závisí predovšetkým na type nervových vlákien a špecifických mozgových systémov, s ktorými sú tieto vlákna pripojené. Pocity každého typu sa prenášajú podľa ich vodivých dráh a typ informácií prenášaných spoločnosťou Neuron je závisí od dráhy, ktorá zahŕňa tento neurón. V akejkoľvek senzorickej prepravnej ceste sa informácie prenášajú z prvého senzorického neurónu (receptor reaguje na vonkajší stimul, ako je dotyk, zápach alebo svetlo) na špecializované neuróny v chrbticom alebo mozgu. Vizuálne informácie sa teda líšia od sluchu len tým, čo sa prenášajú inými vodivými chodníkmi začínajúcimi v sietnici oka a končiace v oblasti mozgu, čo je zodpovedné za vizuálne vnímanie.
Signály odoslané z motorických neurónov mozgu do svalov sú tiež takmer identické so zmyslovými neurónmi z kože do mozgu. Poslúchujú rovnaký princíp "všetko alebo nič", tiež prenášajú intenzitu signálu pomocou frekvencie akčných potenciálov a tiež výsledok signálu závisí len na tom, ktorý neurón je zahrnutý, v ktorom reťazci neuro. Teda rýchla séria potenciálov akcií, ktorá beží na špecifickej vodivé ceste, spôsobuje pohyb vašich prstov, a nie, povedzme, vnímanie viacfarebných svetiel, len preto, že táto cesta je spojená so svalymi rúk a nie s sietnicou.

Všestrannosť potenciálov akcie nie je obmedzená na podobnosť ich prejavov v rôznych neurónoch nachádzajúcich sa v rámci jedného organizmu. Sú to tak isté medzi rôznymi zvieratami, že aj múdre skúsenosti výskumník nie je schopný presne odlíšiť nahrávanie potenciálu nervového vlákna Číny, myš, opice alebo to vedecký líder. Potenciál účinku v rôznych bunkách nie sú identické: malý rozdiel v ich amplitúde a trvanie je stále tam a schválenie "všetky potenciály akcie je také nepresné, ako aj" všetky bougainvilleans sú rovnaké. "
Každý neurón tak prenáša signál cez jeho telo a proces rovnakým spôsobom. Všetky rôzne informácie, ktoré dostávame zo zmyslových neurónov, všetky pohyby, ktoré môžu urobiť naše telo, je výsledkom prenosu jedného typu signálov v rámci neurónov. Stratil "Trifle": Pochopte, čo je signál a ako sa prenáša.

Sme oboznámení s tým, že žijeme živú povahu, vrátane seba, z "non-žijúcich" vecí, vrátane kovových a vysielaní elektrických prúdov. Čím je prekvapujúce si uvedomiť, že v našom telach nie sú kovy jednoducho prítomné - sú potrebné, bez nich nebude schopný existovať. Elektrický prúd - fenomén nie je jednostranný, ale priebežne vznikajúci v sto miliárd neurónov, ktorí prenikli ich procesmi všetky naše telo. Práve teraz môžete zažiť rôzne znaky jeho prítomnosti: skutočnosť, že si uvedomíte, že tento text je výsledkom nespočetných elektrických prúdov. Pocit hladu a potešenie z vône prípravy jedla, vnímanie tohto zápachu, dotyk vetra v jednej rovine do vašej pokožky ... môže byť uvedená nekonečne. A túžba pochopiť, ako sa to všetko vyskytuje, pozostáva tiež z elektrických impulzov vznikajúcich v neurónoch.
Vzhľadom k tomu, účelom tejto kapitoly je posolstvo iba najobecnejších informácií o prechode nervového impulzu, tu je tiež potrebné zvážiť životné prostredie, v ktorom vzniká, tieto podmienky v bunke, ktoré umožňujú vyskytnúť a odovzdať . Preto stojí za to začať s štúdiou premostenia, na ktorom sa udalosti vyvíjajú, a to z Neronu stav odpočinku (stav spiace ['dɔːmənt steɪt]).

Späť v polovici minulého storočia, vedci našli spôsob, ako vytvoriť, v ktorej časti neurónu je elektrický náboj. Na toto použitie voltmeter (voltmeter ['vəultˌmiːtə]) (zariadenie na meranie napätia elektrické pole) S dvoma elektródami. Jedna elektróda je umiestnená vo vnútri neurónu, ktorá je blízko k bunkovej membráne a druhá elektróda je v prostredí neurónu, na druhej strane rovnakej membrány. Voltmeter ukazuje, že z rôznych strán bunkovej membrány existuje elektrické poplatky, negatívne vo vnútri bunky a pozitívne vonku. Existencia takýchto rôznorodých elektrických poplatkov na oboch stranách membrány vytvára elektrické pole, ktoré je dôležitou charakteristikou potenciál. Potenciál, rozprávanie jednoduchým jazykom, je to schopnosť pracovať, napríklad, pracovať na ťahaní nabitej častice z miesta na miesto. Viac negatívnych obvinení na jednej strane a tým pozitívnejšie - na druhej strane membrány, čím silnejšie je elektrické pole, ktoré vytvorili, a silnejšie častice sú schopné tam ťahať. Rozdiel medzi vonkajšími a vnútornými elektrickými poplatkami sa nazýva membránový potenciál (membránový potenciál ['Membreɪn Pə'Tenʃʃl]) odpočinok. Pre neurón je približne 70 mV (Milvolt), to znamená 70 tisíc voltov alebo sedem stotín Volta. Na porovnanie je potenciálny rozdiel v batérii AA 1,5 voltov - 20-krát viac. To znamená, že membránový potenciál zvyšku neurónu je len 20-krát slabší ako medzi svorkami batérie AA - pomerne veľké, ukáže sa. Elektrický potenciál existuje len na membráne av ostatných častiach neutrónovej neutrónov.
Ak píšete presnejšie, membránový potenciál zvyšku Neuron je -70 mV (mínus sedemdesiat milión). Značka mínus znamená len záporný náboj je presne vo vnútri bunky, a nie vonku, a tak sa generované elektrické pole je schopné ťahaním cez membránu vo vnútri bunky pozitívne nabité ióny.
Osoby pri vytváraní membrány potenciálu odpočinku:

1 . V bunková membrána Neuron Existujú kanály, pre ktoré môžu letecké poplatky prechádzať. Zároveň je membrána nie je len pasívnym "oddielom" medzi vnútorným médiom neurónu a okolitou tekutinou: špeciálne proteíny zapustené do membránového mäsa otvorené a zatvorí tieto kanály, a teda membrána ovláda prechod iónov atómy, ktoré majú elektrický náboj. Akumulovanie negatívne nabitých iónov vo vnútri bunky, neurón zvyšuje počet negatívnych obvinení vo vnútri, čo vedie k zvýšeniu pozitívnych nábojov vonku, a tým aj zvyšuje elektrický potenciál. Vzhľadom k tomu, protón má kladný náboj, a volebný elektrón sa získa, potom s nadbytkom protónov, sa získa pozitívne nabitý ión a s nadbytkom elektrónov - negatívne nabitých. Ak chcete podrobnejšie informácie o atómoch a iónoch, môžete sa vrátiť. Je dôležité pochopiť, že membránový potenciál existuje na hranici bunkovej membrány a kvapaliny sú všeobecne vo vnútri a mimo neurónu zostáva elektricky neutrálny. Ióny, pre ktoré membránové prenikanie zostávajú v blízkosti, pretože pozitívne a negatívne obvinenia sa navzájom priťahujú. Výsledkom je, že membrána je tvorená vrstvou "sedí" na IT pozitívnym iónom a vnútri - negatívne. Teda membrána hrá úlohu elektrickej nádoby na oddeľovanie kontajnerov, vo vnútri má elektrické pole. Membrána je preto prirodzeným kondenzátorom.
2 . negatívne nabité proteínyVnútri neurónu v blízkosti vnútorného povrchu membrány. Nabíjanie proteínu vždy zostáva rovnaký a je len súčasťou celkového náboja vnútorného povrchu membrány. Na rozdiel od iónov, proteíny nemôžu opustiť bunku a zadajte ho - pre to sú príliš veľké. Všeobecný poplatok sa líši v závislosti od počtu pozitívnych nabitých iónov umiestnených v blízkosti membrány, ktorej koncentrácia sa môže líšiť cez ich prechod z bunky vonku a zvonku.
3 . Pozitívne nabité ióny draslíka (K +) sa môžu voľne pohybovať medzi vnútorným a vonkajším prostredím, keď je neurón v pokoji. Pohybujú sa neustále otvorenými tečúce kanály draslíka (prietok priechodu draslíka.) Cez ktoré ióny K + môžu prejsť, a nič iné. Tekuté kanály, ktoré nemajú bránu, sa nazývajú, čo znamená otvorené s akýmkoľvek stavom neurónu. Vnútri draselných iónov je oveľa väčšie ako vonku. To sa deje kvôli trvalej práci sodík-draslíkovej pumpy (bude popísané nižšie), takže v stave neurónu iónov K + sa začína pohybovať do vonkajšieho prostredia, pretože koncentrácia tej istej látky sa snaží vyrovnať Vo všeobecnosti. Ak sme v bazéne s vodou v jednom rohu nejakej látky, potom jej koncentrácia v tomto rohu bude veľmi veľká, a v iných častiach bazéna - nula alebo veľmi malé. Avšak, po určitom čase, zistíme, že koncentrácia tejto látky bola vyrovnaná v povodí na úkor brownového hnutia. V tomto prípade hovoria o "čiastočnom tlaku" látky, či už ide o tekutý alebo plyn. Ak sa alkohol vylial do jedného rohu bazéna, potom sa vytvorí veľký rozdiel na koncentrácii alkoholu medzi týmto uhlom a zvyškom bazéna. Dostáva sa čiastočný tlak molekúl alkoholu a budú postupne distribuovať rovnomerne cez bazén, takže čiastočný tlak zmizne, pretože koncentrácia alkoholických molekúl je vyrovnaná všade. Ióny K + nesú pozitívny náboj z neurónu, takže vonku čiastočným tlakom, ktorý je silnejší ako sila príťažlivosti negatívne nabitých proteínov, ak je rozdiel v koncentrácii iónov vo vnútri a mimo bunky dostatočne veľký. Pretože negatívne nabité proteíny zostávajú vo vnútri, je vytvorený záporný náboj na vnútornej strane membrány. Pre jasné pochopenie práce bunkových mechanizmov je dôležité si uvedomiť, že napriek trvalému toku draslíkových iónov z bunky sú vždy väčšie vo vnútri neurónu ako vonku.
4 . Pozitívne nabité sodíkové ióny (NA +) sa nachádzajú na vonkajšej strane membrány a vytvárajú tam pozitívny náboj. Počas fázy buniek sodíkových kanálov NIREP zatvorenéa NA + nemôže ísť dovnútra a ich koncentrácia mimo zvyšovania v dôsledku práce čerpadla sodíka-draslíka, načrtávajú ich z neurónu.
5 . Úloha je negatívne účtovaná ióny chlóru (CL -) a pozitívne nabité ióny vápnika (CA2 +) Ak chcete vytvoriť membránový potenciál, je malý, takže ich správanie zostane za scénami.
Tvorba membrány potenciálu odpočinku Prechádza v dvoch etapách:
Etapa I.. Vytvorí sa malý (-10 mV)) rozdiel potenciálov Čerpadlo sodík-draslík.

Na rozdiel od iných kanálov membrány je vodca draselný sodík schopný prejsť cez seba a ióny sodíka a ióny draslíka. A Na + môže prejsť len z bunky vonku a do + vonku vo vnútri. Jeden cyklus tohto kanála je zahrnutý v 4 krokoch:
1 . "Brána" kanála sodíka-draslík je otvorená len z vnútra membrány a 3 na + tam

2 . Prítomnosť Na + vo vnútri kanála ho ovplyvňuje tak, že môže čiastočne zničiť jednu molekulu ATF (ATP) ( adenosinkosfát), (adenozín trifosfát [ə'dɛnəsiːn trai'fɔsfeɪt]) "Batérie" bunky, pančucha energie a v prípade potreby ho dajte. S takým čiastočným deštrukciou, ktorý spočíva v štiepení od konca molekuly jednej fosfátovej skupiny PO 43-, energia sa rozlišuje, čo sa práve vynaloží na prenos Na + na vonkajší priestor.
3 . Keď sa kanál otvorí, aby sa na + na prepustený, zostal otvorený, a dva ióny spadajú do + - negatívne nabíjačky proteínov zvnútra sú priťahované. Skutočnosť, že v kanáli, ktorá obsahuje tri sodíkové ióny, je umiestnené iba dva ióny draslíka, je celkom logické: atóm draslíka má väčší priemer.
4 . Prítomnosť iónov draslíka teraz, zase, ovplyvňuje kanál, takže externá "brána" je uzavretá a vnútorné otvorí a K + sa prichádza do vnútorného média neurónu.
Tak, sodík-draslík čerpadlo, "výmena" tri sodíkové ióny do dvoch iónov draslíka. Vzhľadom k tomu, elektrický náboj v NA + a K + je rovnaký, ukázalo sa, že z bunky sú odvodené tri pozitívne poplatky, a iba dva padá. Vďaka tomu je znížený vnútorný kladný náboj bunkovej membrány a vonkajšie - zvýšenie. Okrem toho je rozdiel vytvorený v koncentrácii NA + a K + pozdĺž rôznych strán membrány:
\u003d) Mimo bunky sú mnohé sodíkové ióny a vnútri - málo. Súčasne sú sodíkové kanály zatvorené a nedokáže sa dostať späť do Na + bunky a nenechá membránu, pretože priťahuje membránu existujúcu na vnútornej strane.
\u003d) Existuje mnoho iónov draslíka vo vnútri bunky, ale tam je málo z nich vonku, a to vedie k prúdeniu K + z bunkových kanálov otvorených počas údržby neurónu.

Etapa II. Tvorba membrány potenciálu mieru je založená na tomto prúdení iónov draslíka z neurónu. Nižšie uvedený obrázok ukazuje iónové zloženie membrány na začiatku druhého stupňa tvorby potenciálu zvyšku: sada K + a nepriaznivo nabitých proteínov (označených 4-) vo vnútri a membrána naočkovaná mimo Na +. Sťahovanie do vonkajšieho prostredia, ióny draslíka nesú pozitívne návyky z bunky, zatiaľ čo celkový náboj vnútornej membrány sa znižuje. Rovnako ako pozitívne ióny sodíka, tečie z buniek iónov draslíka zostávajú mimo membrány priťahuje vnútorný záporný náboj, a externý kladný náboj membránových záhybov z množstva obvinení NA + a K +. Napriek tečúcim kanálom cez prietokové kanály sú vždy viac vo vnútri buniek iónov draslíka ako vonku.
Vzniká otázka: Prečo ióny draslíka nebudú naďalej prúdiť až do výšky ich množstva vo vnútri bunky a mimo nej sa stáva rovnakým, to znamená, až kým sa čiastočný tlak vytvorený týmito iónmi zmizne? Dôvodom tohto leží v tom, že keď bunky listy, pozitívny náboj sa zvyšuje vonku, a nadbytok záporného náboja sa vytvorí vo vnútri. To znižuje túžbu iónov draslíka opustiť bunku, pretože vonkajší pozitívny náboj ich tlačí a vnútorné negatívne priťahuje. Preto, po chvíli, k + prestane prúdiť, napriek tomu, že v vonkajšom prostredí je ich koncentrácia nižšia ako vo vnútornej: vplyv nábytok na rôznych stranách membrány presahuje silu čiastočného tlaku, to znamená, Prekročí túžbu + distribuovať uniformu v tekutine vnútri a von neurónu. V čase tejto rovnováhy, membránový potenciál neurónu a zastaví asi -70 mV.
Akonáhle je neurón dosiahnutý membránovým potenciálom mieru, je pripravený na vznik a vykonávanie potenciálu činností, ktoré budú diskutované v nasledujúcej cytologickej kapitole.
Zhrnúť: Nerovnomerná distribúcia distribúcie draslíka a iónov sodíka na oboch stranách membrány je spôsobená pôsobením dvoch rivalových síl: a) silou elektrickej príťažlivosti a odpudzovania a b) silou čiastočného tlaku vyplývajúceho z rozdielu v koncentráciách. Práca týchto dvoch rivalových síl pokračuje v podmienkach existencie rozdielne usporiadaných kanálov sodíka, draslíka a draslíka, ktoré pôsobia ako regulátory pôsobenia týchto síl. Draslík kanál tečie, to znamená, že je vždy otvorená v zvyšku neurónu, takže ióny K + môžu ticho chodiť a tu pod vplyvom energie elektromítového odpudzovania / príťažlivosti a pod vplyvom síl čiastočným tlakom, to znamená, že rozdiel v koncentrácii týchto iónov. Sodíkový kanál je vždy uzavretý v stave zvyšku neurónu, takže na + ióny nemusia prechádzať. A nakoniec, kanál sodíka-draslík usporiadaný takým spôsobom, že funguje ako čerpadlo, ktoré s každým cyklom zachytáva tri ióny sodíka smerom von a perámi dvaja ióny draslíka vo vnútri.
Všetok tento dizajn a zabezpečuje výskyt membránového potenciálu zvyšku neurónu: t.j. Štáty, v ktorých sa dosiahli dve veci:
a) Vnútri je záporný poplatok a vonkajšie - pozitívne.
b) Vnútri veľa iónov K +, zamiešané negatívne nabité časti proteínov, a teda dôjde k čiastočnému tlaku draslíka - túžba iónov draslíka na výstup na vyrovnanie koncentrácie.
c) mimo mnohých iónov NA +, ktoré tvoria čiastočne pár s iónmi CL. A teda dochádza k čiastočnému tlaku sodného - túžba sodíkových iónov vstupujú do bunky, aby sa koncentrácia vyrovnala.
V dôsledku práce čerpadla sodného draslíka dostaneme tri sily, ktoré existujú na membráne: výkon elektrického poľa a pevnosť dvoch čiastkových tlakov. Tieto sily a začať pracovať, keď neurón opustí stav odpočinku.
Otvorenie histórie
V roku 1902, Julius Bernstein zdôraznil hypotézu, podľa ktorej bunková membrána prechádza do iónov buniek na + a hromadia sa v cytoplazme. Výpočet veľkosti potenciálu starostlivosti podľa Nernstovej rovnice pre potenciálnu elektródu uspokojivo sa zhodovala s nameraným potenciálom medzi svarkoplazmou svalmi a environmentálnyČo bolo asi 70 mv.
Podľa Y. Bernstein teórie, keď je bunka vzrušený, jeho membrána je poškodená a ióny K + unikajú z bunky v koncentračnom gradiente, až kým sa membránový potenciál nechajú rovný nule. Membrána potom obnovuje svoju integritu a potenciál sa vracia na úroveň potenciálu odpočinku. Toto vyhlásenie o potenciáli o potenciáli bolo vyvrátilo Hodgkin a Huxley v roku 1939.
Bernsteinova teória týkajúca sa potenciálu starostlivosti potvrdila Kenneth Stewart Cole (Kenneth Stewart Cole), niekedy jeho iniciály sa mylne napísané ako K.C. Cole, pretože jeho prezývka, Casey ("Kacy"). PP a PD sú znázornené na známe ilustráciu Coul a Curtis, 1939. Tento obrázok sa stal znakom membránovej biofyziky skupiny biofyzikálnej spoločnosti (pozri ilustráciu).
Všeobecné ustanovenia
Aby bola membrána podporovaná potenciálnym rozdielom, je potrebné, aby určitý rozdiel v koncentrácii rôznych iónov vo vnútri a mimo bunky.
Koncentrácia iónov v bunke kostrovej svalovej a v extracelulárnom médiu
Potenciál ľudí pre väčšinu neurónov je asi -60 mV - -70 mV. V bunkách non-profesionálnych tkanív má membrána tiež rozdiel v potenciáloch, odlišný pre bunky rôznych tkanív a organizmov.
Vytvorenie potenciálu odpočinku
PP je vytvorený v dvoch etapách.
Prvé štádium: Vytvorenie menšieho (-10 mV) negativity vo vnútri bunky v dôsledku nerovnakej asymetrickej výmeny NA + na K + v pomere 3: 2. V dôsledku toho bunka opustí pozitívnejšie návyky s sodíkom, ako sa vráti s draslíkom. Takýto znak práce čerpadla sodíka-draslíka, ktorý vykonáva výmenu týchto iónov cez membránu s nákladmi na energiu ATP, poskytuje svoju elektrickú energiu.
Výsledky činností membránových iónových čerpadiel v prvej fáze tvorby PP sú nasledovné: \\ t
1. Nedostatok iónov sodného (Na +) v bunke.
2. Prebytočné ióny draslíka (K +) v bunke.
3. Vzhľad na membráne slabého elektrického potenciálu (-10 mV).
Druhá fáza: Tvorba významného (-60 mV) negativity vo vnútri bunky kvôli úniku z neho cez membránu K + iónov. Kaliya K + ióny opúšťajú klietku a vykonávajú pozitívne poplatky s nimi, prinášajúc na -70 mV.
Potenciál starostlivosti o membrán je teda nedostatok pozitívnych elektrických nábojov vo vnútri bunky, ku ktorej dochádza v dôsledku úniku z neho pozitívnych iónov draslíka a elektrickým účinkom čerpadla sodíka.
pozri tiež
Poznámky
Spojenie
Dudel J., Roegg J., Schmidt R., atď. Ľudská fyziológia: V 3 zväzkoch. Za. Od angličtiny / v blízkosti Ed R. Schmidt a mesto TEVS. - 3. - m.: Mir, 2007. - T. 1. - 323 s chorý. z. - 1500 kópií. - ISBN 5-03-000575-3.
Nadácia Wikimedia. 2010.
Sledujte, čo je "potenciál mieru" v iných slovníkoch:
Potenciál, elektrický potenciál medzi vnútorným a vonkajším médiom sa vyskytuje na jeho membráne; Neuróny a svalové bunky dosiahne hodnotu 0,05 0,09 V; Vyskytuje sa kvôli nerovnomernej distribúcii a akumulácii iónov v rôznych ... Encyklopedický slovník
Membránový potenciál mieru, potenciálny rozdiel existujúci v živých bunkách v stave fyziky. Medzi ich cytoplazmou a extracelulárnou tekutinou. Nervové a svalnaté bunky P. p. Rôzne zvyčajne v rozsahu 60,90 mV a interné. Strana ...
potenciálny odpočinok - Stres odpočinku - [Ya.n. Luginsky, M.S.FESI ZHILINSKAYA, YU.S. KABIROV. Anglický Ruský slovník pre elektrotechnické a elektrické energetické priemysel, Moskva, 1999] elektrické zariadenia Témy, základné koncepty Synonymá Sixers En ODSTRÁNOVAŤ POTREBUJÚCICH ... Technický adresár prekladateľa
potenciálny odpočinok - Zvyšok (ING) Potenciálny potenciálny potenciál existujúci medzi médiom, v ktorom je bunka umiestnená a jej obsah ... Vysvetľujúci angličtina-Ruský slovník o nanotechnológii. - M.
Potenciálny odpočinok - potenciál neaktívneho neurónu. To sa nazýva aj membránový potenciál ... Psychológia pocitov: Slovník
potenciálny odpočinok - potenciálny rozdiel medzi obsahom bunky a extracelulárnou tekutinou. V nervových bunkách p.p. Zúčastňuje sa na udržiavaní pripravenosti bunky k vzrušeniu. * * * * Membránový bioelektrický potenciál (asi 70mb) v nervovej bunke sa nachádza v ... ... Encyklopedický slovník psychológie a pedagogiky
Potenciálny odpočinok - - Rozdiel elektrických obvinení medzi vonkajším a vnútorným povrchom membrány v stave fyziologickej strany bunky, zaregistrovaný pred začiatkom dráždivého ... Slovník termínov vo fyziológii poľnohospodárskych zvierat
Membránový potenciál zaregistrovaný pred začiatkom dráždivého ... Veľký lekársky slovník
- (fyziologický) potenciálny rozdiel medzi obsahom bunky (vlákniny) a extracelulárnou kvapalinou; Potenciálny skok je lokalizovaný na povrchovej membráne, s ním je vnútorná strana nabitá elektronegatívne s ohľadom na ... ... Veľká sovietska encyklopédia
Rýchle oscilácie (hrot) membránového potenciálu vznikajúceho z excitácie nervových, svalov, ne-ryy žliaz a pestovania, buniek; Elektrické. Signál poskytuje rýchly prenos informácií v tele. Oberys pravidlo "všetko alebo nič" ... ... Biologický encyklopedický slovník
Knihy
- 100 spôsobov, ako zmeniť život. Časť 1, Parthenteva Larisa. O knihe Zbierka inšpiratívnych príbehov o tom, ako zmeniť život pre lepšie, od osoby, ktorá sa podarilo nasadiť svoj vlastný život o 180 stupňov. Táto kniha sa narodila od týždenného okruhu ...
Bolo zistené, že najdôležitejšie ióny, ktoré určujú potenciály membránových buniek, sú anorganické ióny K +, NA +, SG, ako aj v niektorých prípadoch CA2 +. Je dobre známe, že koncentrácie týchto iónov v cytoplazme a v intercelulárnej tekutine sa líšia desiatkami.
Z tabuľky. 11.1 Je možné vidieť, že koncentrácia iónov na + vo vnútri bunky je 40-60-krát vyššia ako v intercelulárnej tekutine, zatiaľ čo pre NA + a SG je distribúcia koncentrácií naopak. Nerovná distribúcia koncentrácií týchto iónov na oboch stranách membrány je zabezpečená oboma rôznou permeabilitou a silným elektrickým membránovým poľom, ktorý je určený jeho zvyšným potenciálom.
V skutočnosti, v stave odpočinku, celkový tok iónov cez membránu je nula, a potom z rovnice non-rýnie - doska z toho vyplýva
Tak, v pokojovej koncentrácii - a
elektrický potenciál - na membráne zameranom
proti sebe, a preto v pokojovej bunke vysoký a konštantný rozdiel koncentrácií hlavných iónov zaisťuje, že bunky elektrickej napätej bunky na membráne sa nazýva, ktoré sa nazývajú equilibrium membránový potenciál.
Na druhej strane, špičkový potenciál, ktorý sa vyskytuje na membráne zabraňuje výstupu iónu z bunky K + a nadmerného vstupu do IT SG, čím sa udržiavajú koncentrácie gradienty na membráne.
Úplný výraz pre membránový potenciál, ktorý berie do úvahy difúzne toky týchto troch typov iónov, získal Goldman, Hodgkin a Katz:
kde R k P Na, p C1 - permeabilita membrány pre zodpovedajúce ióny.
Rovnica (11.3) s vysokou presnosťou určuje membránové potenciály odpočinku rôzne bunky. Z toho vyplýva, že pre membránový potenciál odpočinku nie je dôležitý absolútne hodnoty Priepustnosť membrány pre rôzne ióny a ich vzťah, pretože rozdelenie oboch častí frakcie pod znakom logaritmu, napríklad na R K, prejdeme k relatívnej permeabilite iónov.
V prípadoch, keď je priepustnosť jedného z týchto iónov výrazne väčšia ako iné, rovnica (11.3) vstupuje do Nernstovej rovnice (11.1) na tento ion.
Z tabuľky. 11.1 Je možné vidieť, že membránový potenciál buniek buniek je blízky potenciálu NernSTA pre ióny K + a CV, ale je z neho výrazne odlišné od neho NA +. Toto je dôkaz
0 Skutočnosť, že v zvyšku membrány je dobre priepustná pre ióny K + a SG, zatiaľ čo pre ióny + jeho priepustnosť je veľmi nízka.
Napriek tomu, že rovnovážny potenciál NernSTA pre SG je najbližšie k potenciálu zvyšku klietky, druhý má prevažne draslík. Je to spôsobené tým, že vysoká intracelulárna koncentrácia K + sa nemôže významne znížiť, pretože ióny K + by mali vyvážiť vo vnútri objemového negatívneho aniónu buniek. Intracelulárne anióny sú väčšinou veľké organických molekúl (proteíny, zvyšky organických kyselín IT.P.), ktoré nemôžu prejsť cez kanály v bunkovej membráne. Koncentrácia týchto aniónov v bunke je takmer konštantná a ich celkový záporný náboj zabraňuje významu výstupu draslíka z bunky, pričom sa udržiava jeho vysoká intracelulárna koncentrácia spolu s čerpadlom Na-K. Hlavná úloha v počiatočnom zariadení v rámci bunky s vysokou koncentráciou iónov draslíka a nízkych koncentrácií sodíka patrí k N-K-čerpadlom.
Distribúcia iónov C1 je stanovená v súlade s membránovým potenciálom, pretože neexistujú žiadne špeciálne mechanizmy na udržanie koncentrácie SG v bunke. Preto je v dôsledku negatívneho nabitia chlóru, jeho distribúcia je opačná s ohľadom na distribúciu draslíka na membráne (pozri tabuľku 11.1). Difúzia koncentrácie K + z bunky a C1 do bunky je teda prakticky vyrovnaná membránovým potenciálom zvyšku bunky.
Pokiaľ ide o NA +, potom je jej difúzia nasmerovaná do bunky pod pôsobením koncentračného gradientu a membránového elektrického poľa a vstup Na + do bunky je obmedzený na bližiacu iba permeabilitu membrány pre sodík (sodík kanály sú zatvorené). Skutočne, Hodgkin a Katz experimentálne zistili, že v stave zvyšku permeability axónovej membrány SQUID pre K +, NA + a SG sa vzťahujú na 1: 0,04: 0,45. Tak, v stave zvyšku, bunková membrána je len priepustná pre NA +, a pre SG prenikne takmer tak dobre ako pre K +. V nervových bunkách je permeabilita pre SG zvyčajne nižšia ako pre K +, ale vo svalovej vláknach je priepustnosť pre SG dokonca trochu prevláda.
Napriek malej priepustnosti bunkovej membrány pre samotné Na + existuje, aj keď veľmi malý, pasívny prenos NA + do bunky. Tento prúd Na + by musel viesť k zníženiu potenciálneho rozdielu na membráne a na výstup na + z bunky, ktorý by v konečnom dôsledku veril, že zarovnanie koncentrácií NA + a K + na oboch stranách membrány. To sa nevyskytuje v dôsledku práce NA + - K + -pasos, ktorý kompenzuje únikové prúdy NA + a K + a udržiavanie normálne hodnoty Intracelulárne koncentrácie týchto iónov, a preto normálne množstvo potenciálu bunkového odpočinku.
Pre väčšinu buniek je membránový odpočinkový potenciál (-beyond) - (- yuo) mV. Na prvý pohľad sa môže zdať, že ide o malú hodnotu, ale je potrebné zvážiť, že membránová hrúbka je tiež malá (8-10 nm), takže silná sila elektrického poľa v bunkovej membráne je enormný a je približne 10 miliónov Volts na 1 m (alebo 100 m2 na 1 cm):
Napríklad vzduch, nevydrží takáto silu elektrického poľa (elektrický rozpad vo vzduchu sa vyskytuje pri 30 kV / cm) a membrána môže vydržať. Toto je normálny stav svojej aktivity, pretože je presne takáto elektrické pole na udržanie rozdielu v koncentráciách sodných iónov, draslíka a chlóru na membráne.
Veľkosť potenciálu mieru, odlišný v bunkách, môže zmeniť, keď podmienky menia svoje živobytie. Porušenie bioenergie procesov v bunke, sprevádzané poklesom intracelulárnej hladiny makro-ergických zlúčenín (najmä ATP), primárne eliminuje špičkový potenciálny komponent spojený s prácou MA + -K + -atf- ASE.
Príčiny poškodenia buniek zvyčajne zvyšujú priepustnosť bunkových membrán, v dôsledku čoho sa rozdiely v permeabilite membrány pre draslíka a ióny sodíka znížia; Potenciál odpočinku sa znižuje, čo môže spôsobiť porušenie viacerých bunkových funkcií, ako je excitabilita.
- Pretože intracelulárna koncentrácia draslíka je podopretá takmer konštantnou, dokonca aj relatívne malé zmeny v extracelulárnej koncentrácii môžu mať výrazný účinok na potenciálne vnímanie a bunkovú aktivitu. Podobné zmeny koncentrácie K "v krvnej plazme sa vyskytujú v niektorých patológiách (napr.