Мембранна різниця потенціалів. Основні властивості нервової клітини. Клітинна мембрана як електричний ланцюг
Стаття на конкурс «біо / мовляв / текст»: Потенціал спокою - це важливе явище в житті всіх клітин організму, і важливо знати, як він формується. Однак це складний динамічний процес, важкий для сприйняття цілком, особливо для студентів молодших курсів (біологічних, медичних і психологічних спеціальностей) і непідготовлених читачів. Втім, при розгляді по пунктам, цілком можливо зрозуміти його основні деталі і етапи. В роботі вводиться поняття потенціалу спокою і виділяються основні етапи його формування з використанням образних метафор, які допомагають зрозуміти і запам'ятати молекулярні механізми формування потенціалу спокою.
Мембранні транспортні структури - натрій-калієві насоси - створюють передумови для виникнення потенціалу спокою. Передумови ці - різниця в концентрації іонів на внутрішній і зовнішній сторонах клітинної мембрани. Окремо проявляє себе різницю концентрації по натрію і різниця концентрації по калію. Спроба іонів калію (K +) вирівняти свою концентрацію по обидві сторони мембрани призводить до його витоку з клітки і втрати разом з ними позитивних електричних зарядів, За рахунок чого значно посилюється загальний негативний заряд внутрішньої поверхні клітини. Ця «калієва» негативність становить більшу частину потенціалу спокою (-60 мВ в середньому), а меншу його частину (-10 мВ) становить «обмінна» негативно, викликана Електрогене самого іонного насоса-обмінника.
Давайте розбиратися докладніше.
Навіщо нам потрібно знати, що таке потенціал спокою і як він виникає?
Ви знаєте, що таке «тварина електрику»? Звідки в організмі беруться «біоструми»? як жива клітина, Що знаходиться у водному середовищі, може перетворитися в «електричну батарейку» і чому вона моментально нерозряджаються?
На ці питання можна відповісти тільки в тому випадку, якщо дізнатися, як клітина створює собі різницю електричних потенціалів (потенціал спокою) на мембрані.
Цілком очевидно, що для розуміння того, як працює нервова система, необхідно спочатку розібратися, як працює її окрема нервова клітина - нейрон. Головне, що лежить в основі роботи нейрона - це переміщення електричних зарядів через його мембрану і поява внаслідок цього на мембрані електричних потенціалів. Можна сказати, що нейрон, готуючись до своєї нервової роботі, спочатку запасає енергію в електронній формі, а потім використовує її в процесі проведення і передачі нервового збудження.
Таким чином, наш найперший крок до вивчення роботи нервової системи - це зрозуміти, яким чином з'являється електричний потенціал на мембрані нервових клітин. Цим ми і займемося, і назвемо цей процес формуванням потенціалу спокою.
Визначення поняття «потенціал спокою»
У нормі, коли нервова клітина знаходиться в фізіологічному спокої і готова до роботи, у неї вже відбувся перерозподіл електричних зарядів між внутрішньою і зовнішньою сторонами мембрани. За рахунок цього виникло електричне поле, і на мембрані з'явився електричний потенціал - мембранний потенціал спокою.
Таким чином, мембрана виявляється поляризованої. Це означає, що вона має різний електричний потенціал зовнішньої і внутрішньої поверхонь. Різниця між цими потенціалами цілком можливо зареєструвати.
У цьому можна переконатися, якщо ввести всередину клітини мікроелектрод, з'єднаний з реєструє установкою. Як тільки електрод потрапляє всередину клітини, він миттєво набуває певний постійний електронегативний потенціал по відношенню до електрода, розташованого в навколишньому клітку рідини. Величина внутрішньоклітинного електричного потенціалу у нервових клітин і волокон, наприклад, гігантських нервових волокон кальмара, в спокої становить близько -70 мВ. Цю величину називають мембранним потенціалом спокою (МПП). У всіх точках аксоплазми цей потенціал практично однаковий.
Ноздрачов А.Д. та ін. Почала фізіології.
Ще трохи фізики. Макроскопічні фізичні тіла, як правило, електрично нейтральні, тобто в них в рівних кількостях містяться як позитивні, так і негативні заряди. Зарядити тіло можна, створивши в ньому надлишок заряджених частинок одного виду, наприклад, тертям про інше тіло, в якому при цьому утворюється надлишок зарядів протилежного виду. З огляду на наявність елементарного заряду ( e), Повний електричний заряд будь-якого тіла можна уявити як q \u003d ± N × e, Де N - ціле число.
потенціал спокою - це різниця електричних потенціалів, наявних на внутрішній і зовнішній сторонах мембрани, коли клітина знаходиться в стані фізіологічного спокою. Його величина вимірюється зсередини клітини, вона негативна і становить в середньому -70 мВ (мілівольт), хоча в різних клітинах може бути різною: від -35 мВ до -90 мВ.
Важливо враховувати, що в нервовій системі електричні заряди представлені не електронами, як у звичайних металевих дротах, а іонами - хімічними частками, мають електричний заряд. І взагалі у водних розчинах у вигляді електричного струму переміщаються не електронний, а іони. Тому всі електричні струми в клітинах і навколишнього їхньому середовищі - це іонні струми.
Отже, зсередини клітина в спокої заряджена негативно, а зовні - позитивно. Це властиво всім живим клітинам, за винятком, хіба що, еритроцитів, які, навпаки, заряджені негативно зовні. Якщо говорити конкретніше, то виходить, що зовні навколо клітини будуть переважати позитивні іони (катіони Na \u200b\u200b+ і K +), а всередині - негативні іони (аніони органічних кислот, які не здатні вільно переміщатися через мембрану, як Na + і K +).
Тепер нам всього лише залишилося пояснити, яким же чином все вийшло саме так. Хоча, звичайно, неприємно усвідомлювати, що всі наші клітини крім еритроцитів тільки зовні виглядають позитивними, а всередині вони - негативні.
Термін «негативність», який ми будемо застосовувати для характеристики електричного потенціалу всередині клітини, стане в нагоді нам для простоти пояснення змін рівня потенціалу спокою. За цим терміном приємно те, що інтуїтивно зрозуміло наступне: чим більше негативність всередині клітини - тим нижче в негативну сторону від нуля зміщений потенціал, а чим менше негативність - тим ближче негативний потенціал до нуля. Це набагато простіше зрозуміти, чим кожен раз розбиратися в тому, що ж саме означає вираз «потенціал зростає» - зростання за абсолютним значенням (або «по модулю») буде означати зсув потенціалу спокою вниз від нуля, а просто «зростання» - зміщення потенціалу вгору до нуля. Термін «негативність" не створює подібних проблем неоднозначності розуміння.
Сутність формування потенціалу спокою
Спробуємо розібратися, звідки береться електричний заряд нервових клітин, хоча їх ніхто не тре, як це роблять фізики в своїх дослідах з електричними зарядами.
Тут дослідника і студента чекає одна з логічних пасток: внутрішня негативність клітини виникає не через появи зайвих негативних частинок (Аніонів), а, навпаки, через втрати деякої кількості позитивних частинок (Катіонів)!
Так куди ж діваються з клітки позитивно заряджені частинки? Нагадаю, що це покинувши клітку і скупчилися зовні іони натрію - Na + - і калію - K +.
Головний секрет появи отріцательності всередині клітини
Відразу відкриємо цей секрет і скажемо, що клітина позбавляється частини своїх позитивних частинок і заряджається негативно за рахунок двох процесів:
- спочатку вона обмінює «свій» натрій на «чужій» калій (так-так, одні позитивні іони на інші, такі ж позитивні);
- потім з неї відбувається витік цих «наміняти» позитивних іонів калію, разом з якими з клітки витікають позитивні заряди.
Ці два процеси нам і треба пояснити.
Перший етап створення внутрішньої отріцательності: обмін Na + на K +
У мембрані нервової клітини постійно працюють білкові насоси-обмінники(Аденозінтріфосфатази, або Na + / K + -АТФази), вбудовані в мембрану. Вони змінюють «власний» натрій клітини на зовнішній «чужий» калій.
Але ж при обміні одного позитивного заряду (Na +) на інший такий же позитивний заряд (K +) ніякого дефіциту позитивних зарядів в клітці виникати не може! Правильно. Але, тим не менш, через це обміну в клітині залишається дуже мало іонів натрію, тому що вони майже всі пішли назовні. І в той же час клітина переповнюється іонами калію, які в неї накачали молекулярні насоси. Якби ми могли спробувати на смак цитоплазму клітини, ми б помітили, що в результаті роботи насосів-обмінників вона перетворилася з солоною в гірко-солоно-кислий, тому що солоний смак хлориду натрію змінився складним смаком досить-таки концентрованого розчину хлориду калію. У клітці концентрація калію досягає 0,4 моль / л. Розчини хлориду калію в межах 0,009-0,02 моль / л мають солодкий смак, 0,03-0,04 - гіркий, 0,05-0,1 - гірко-солоний, а починаючи з 0,2 і вище - складний смак , що складається з солоного, гіркого і кислого.
Важливо тут те, що обмін натрію на калій - нерівний. За кожні віддані кліткою три іона натрію вона отримує все два іона калію. Це призводить до втрати одного позитивного заряду при кожному акті іонного обміну. Так що вже на цьому етапі за рахунок нерівноцінного обміну клітина втрачає більше «плюсів», ніж отримує взамін. В електричному вираженні це становить приблизно -10 мВ отріцательності всередині клітини. (Але пам'ятайте, що нам треба ще знайти пояснення для решти -60 мВ!)
Щоб легше було запам'ятати роботу насосів-обмінників, образно можна висловитися так: «Клітка любить калій!»Тому клітка і затягує калій до себе, незважаючи на те, що його і так в ній повно. І тому вона невигідно обмінює його на натрій, віддаючи 3 іона натрію за 2 іона калію. І тому вона витрачає на цей обмін енергію АТФ. І як витрачає! До 70% всіх енерговитрат нейрона може йти на роботу натрій-калієвих насосів. (Ось що робить любов, нехай вона навіть і не справжня!)
До речі, цікаво, що клітина не народжується з готовим потенціалом спокою. Їй його ще треба створити. Наприклад, при диференціюванні і злиття міобластів потенціал їх мембрани змінюється від -10 до -70 мВ, тобто їх мембрана стає більш негативною - поляризується в процесі диференціювання. А в експериментах на мультіпотентних мезенхімальних стромальних клітинах кісткового мозку людини штучна деполяризация, що протидіє потенціалу спокою і зменшує негативність клітин, навіть інгібувати (гнітила) диференціювання клітин.
Образно кажучи, можна висловитися так: Створюючи потенціал спокою, клітка «заряджається любов'ю». Це любов до двох речей:
- любов клітини до калію (тому клітина насильно затягує його до себе);
- любов калію до свободи (тому калій залишає захопила його клітку).
Механізм насичення клітини калієм ми вже пояснили (це робота насосів-обмінників), а механізм догляду калію з клітки пояснимо нижче, коли перейдемо до опису другого етапу створення внутрішньоклітинної отріцательності. Отже, результат діяльності мембранних іонних насосів-обмінників на першому етапі формування потенціалу спокою такий:
- Дефіцит натрію (Na +) в клітці.
- Надлишок калію (K +) в клітці.
- Поява на мембрані слабкого електричного потенціалу (-10 мВ).
Можна сказати так: на першому етапі іонні насоси мембрани створюють різницю концентрацій іонів, або градієнт (перепад) концентрації, між внутрішньоклітинної і позаклітинної середовищем.
Другий етап створення отріцательності: витік іонів K + з клітини
Отже, що починається в клітці після того, як з іонами попрацюють її мембранні натрій-калієві насоси-обмінники?
Через утворився дефіциту натрію всередині клітини цей іон при кожному зручному випадку норовить спрямуватися всередину: Розчинені речовини завжди прагнуть вирівняти свою концентрацію в усьому об'ємі розчину. Але це у натрію виходить погано, оскільки іонні натрієві канали зазвичай закриті і відкриваються тільки за певних умов: під впливом спеціальних речовин (трансмиттеров) або при зменшенні отріцательності в клітці (деполяризаціїмембрани).
У той же час в клітці є надлишок іонів калію в порівнянні з зовнішнім середовищем - бо насоси мембрани насильно накачали його в клітку. І він, теж прагнучи зрівняти свою концентрацію всередині і зовні, норовить, навпаки, вийти з клітки. І це у нього виходить!
Іони калію K + залишають клітку під дією хімічного градієнта їх концентрації по різні боки мембрани (мембрана значно більш проникна для K +, ніж для Na +) і забирають з собою позитивні заряди. Через це всередині клітини наростає негативність.
Тут ще важливо зрозуміти те, що іони натрію і калію як би «не помічають» один одного, вони реагують тільки «на самих себе». Тобто натрій реагує на концентрацію натрію ж, але «не звертає уваги» на те, скільки навколо калію. І навпаки, калій реагує тільки на концентрацію калію і «не помічає» натрій. Виходить, що для розуміння поведінки іонів треба окремо розглядати концентрації іонів натрію і калію. Тобто треба окремо порівняти концентрацію по натрію всередині і зовні клітини і окремо - концентрацію по калію всередині і зовні клітини, але не має сенсу порівнювати натрій з калієм, як це, буває, робиться в підручниках.
Згідно із законом вирівнювання хімічних концентрацій, який діє в розчинах, натрій «хоче» зовні увійти в клітку; туди ж його тягне і електрична сила (як ми пам'ятаємо, цитоплазма заряджена негативно). Хотіти щось він хоче, але не може, так як мембрана в звичайному стані погано його пропускає. Натрієві іонні канали, наявні в мембрані, в нормі закриті. Якщо все ж його заходить трошки, то клітина відразу ж обмінює його на зовнішній калій за допомогою своїх натрій-калієвих насосів-обмінників. Виходить, що іони натрію проходять через клітку як би транзитом і не затримуються в ній. Тому натрій в нейронах завжди в дефіциті.
А ось калій якраз може легко виходити з клітки назовні! У клітці його повно, і вона його втримати не може. Він виходить назовні через особливі канали в мембрані - «калієві канали витоку», які в нормі відкриті і випускають калій.
До + -канали витоку постійно відкриті при нормальних значеннях мембранного потенціалу спокою і виявляють вибухи активності при зрушеннях мембранного потенціалу, які тривають кілька хвилин і спостерігаються при всіх значеннях потенціалу. Посилення До + -Ток витоку веде до гіперполяризації мембрани, тоді як їх придушення - до деполяризації. ... Однак, існування канального механізму, відповідального за струми витоку, довгий час залишалося під питанням. Тільки зараз стало ясно, що калієва витік - це струм через спеціальні калієві канали.
Зефиров А.Л. і Сітдікова Г.Ф. Іонні канали збудливою клітини (структура, функція, патологія).
Від хімічного - до електричного
А тепер - ще раз найголовніше. Ми повинні усвідомлено перейти від руху хімічних частинок до руху електричних зарядів.
Калій (K +) позитивно заряджений, і тому він, коли виходить з клітки, виносить з неї не тільки самого себе, а й позитивний заряд. За ним зсередини клітини до мембрани тягнуться «мінуси» - негативні заряди. Але вони не можуть проникнути через мембрану - на відміну від іонів калію - тому що для них немає відповідних іонних каналів, і мембрана їх не пропускає. Пам'ятайте про решту непоясненим нами -60 мВ отріцательності? Це і є та сама частина мембранного потенціалу спокою, яку створює витік іонів калію з клітки! І це - велика частина потенціалу спокою.
Для цієї складової частини потенціалу спокою є навіть спеціальну назву - концентраційний потенціал. концентраційний потенціал - це частина потенціалу спокою, створена дефіцитом позитивних зарядів всередині клітини, що утворився за рахунок витоку з неї позитивних іонів калію.
Ну, а тепер трохи фізики, хімії та математики для любителів точності.
Електричні сили пов'язані з хімічними за рівнянням Гольдмана. Його окремим випадком є \u200b\u200bбільш просте рівняння Нернста, по формулі якого можна розрахувати трансмембранну диффузионную різниця потенціалів на основі різної концентрації іонів одного виду по різні боки мембрани. Так, знаючи концентрацію іонів калію зовні і всередині клітини, можна розрахувати калієвий рівноважний потенціал E K:

де Едо - рівноважний потенціал, R - газова постійна, Т - абсолютна температура, F - постійна Фарадея, К + зовн і K + внутр - концентрації іонів К + зовні і всередині клітини, відповідно. За формулою видно, що для розрахунку потенціалу між собою порівнюються концентрації іонів одного виду - K +.
Більш точно підсумкова величина сумарного дифузійного потенціалу, який створюється витоком декількох видів іонів, розраховується за формулою Гольдмана-Ходжкіна-Катца. У ній враховано, що потенціал спокою залежить від трьох чинників: (1) полярності електричного заряду кожного іона; (2) проникності мембрани Р для кожного іона; (3) [концентрацій відповідних іонів] всередині (внутр) і зовні мембрани (зовн). Для мембрани аксона кальмара в спокої відношення провідностей Р K: PNa :P Cl \u003d 1: 0,04: 0,45.
висновок
Отже, поті нціал спокою складається з двох частин:
- -10 мВ, Які виходять від «несиметричною» роботи мембранного насоса-обмінника (адже він більше викачує з клітини позитивних зарядів (Na +), ніж закачує назад з калієм).
- Друга частина - це весь час витікає з клітки калій, що відносить позитивні заряди. Його внесок - основний: -60 мВ. В сумі це і дає шукані -70 мВ.
Що цікаво, калій перестане виходити з клітки (точніше, його вхід і вихід зрівнюються) тільки при рівні отріцательності клітини -90 мВ. В цьому випадку зрівняються хімічні та електричні сили, які проштовхують калій через мембрану, але напрямні його в протилежні сторони. Але цьому заважає постійно підтікає в клітку натрій, який несе з собою позитивні заряди і зменшує негативно, за яку «бореться» калій. І в підсумку в клітці підтримується рівноважний стан на рівні -70 мВ.
Ось тепер мембранний потенціал спокою остаточно сформований.
Схема роботи Na + / K + -АТФази наочно ілюструє «несиметричний» обмін Na + на K +: викачування надлишкового «плюса» в кожному циклі роботи ферменту призводить до негативного заряджені внутрішньої поверхні мембрани. Чого в цьому ролику не сказано, так це того, що АТФаза відповідальна за менш ніж 20% потенціалу спокою (-10 мВ): залишилася «негативність» (-60 мВ) з'являється за рахунок виходу з клітини через «калієві канали витоку» іонів K +, які прагнуть вирівняти свою концентрацію всередині клітини і поза нею.
література
- Jacqueline Fischer-Lougheed, Jian-Hui Liu, Estelle Espinos, David Mordasini, Charles R. Bader, et. al .. (2001). Human Myoblast Fusion Requires Expression of Functional Inward Rectifier Kir2.1 Channels. J Cell Biol. 153 , 677-686;
- Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. (1998). Role of an inward rectifier K + current and of hyperpolarization in human myoblast fusion. J. Physiol. 510 , 467–476;
- Sarah Sundelacruz, Michael Levin, David L. Kaplan. (2008). Membrane Potential Controls Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells. PLoS ONE. 3 , E3737;
- Павловська М.В. і Мамикіна А.І. Електростатика. Діелектрики і провідники в електричному полі. Постійний струм / електронний посібник по загальному курсу фізики. СПб: Санкт-Петербурзький державний електротехнічний університет;
- Ноздрачов А.Д., Баженов Ю.І., Баранникова І.А., Батуев А.С. та ін. Почала фізіології: Підручник для вузів / Під ред. акад. А.Д. Ноздрачов. СПб: Лань, 2001. - тисячі вісімдесят вісім с .;
- Макаров А.М. і Луньова Л.А. Основи електромагнетизму / Фізика в технічному університеті. Т. 3;
- Зефиров А.Л. і Сітдікова Г.Ф. Іонні канали збудливою клітини (структура, функція, патологія). Казань: Арт-кафе, 2010. - 271 с .;
- Родина Т.Г. Сенсорний аналіз продовольчих товарів. Підручник для студентів вузів. М .: Академія, 2004. - 208 с .;
- Кольман Я. і Рем К.-Г. Наочна біохімія. М .: Світ, 2004. - 469 с .;
- Шульговскій В.В. Основи нейрофізіології: Навчальний посібник для студентів вищих навчальних закладів. М .: Аспект Пресс, 2000. - 277 с ..
Будь-яка жива клітина покрита напівпроникною мембраною, через яку здійснюється пасивне рух і активний виборчий транспорт позитивно і негативно заряджених іонів. Завдяки цьому переносу між зовнішньою і внутрішньою поверхнею мембрани є різниця електричного заряду (потенціалів) - мембранний потенціал. Існує три відрізняються один від одного прояву мембранного потенціалу - мембранний потенціал спокою, місцевий потенціал, або локальний відповідь, і потенціал дії.
Якщо на клітину не діють зовнішні подразники, то мембранний потенціал довго зберігається постійним. Мембранний потенціал такої покоїться клітини називається мембранним потенціалом спокою. Для зовнішньої поверхні мембрани клітини потенціал спокою завжди позитивний, а для внутрішньої поверхні клітинної мембрани завжди негативний. Прийнято вимірювати потенціал спокою на внутрішній поверхні мембрани, тому що іонний склад цитоплазми клітини більш стабільний, ніж міжклітинної рідини. Величина потенціалу спокою відносно постійна для кожного типу клітин. Для поперечносмугастих м'язових клітин вона становить від -50 до -90 мВ, а для нервових клітин від -50 до -80 мВ.
Причинами виникнення потенціалу спокою є різна концентрація катіонів та аніонів зовні і всередині клітини, а також виборча проникність для них клітинної мембрани. Цитоплазма спочиває нервової і м'язової клітини містить приблизно в 30-50 разів більше катіонів калію, в 5-15 разів менше катіонів натрію і в 10-50 разів менше аніонів хлору, ніж позаклітинна рідина.
У стані спокою практично всі натрієві канали мембрани клітини закриті, а більшість калієвих каналів відкрито. Всякий раз, коли іони калію наштовхуються на відкритий канал, вони проходять через мембрану. Оскільки всередині клітини іонів калію набагато більше, то осмотическая сила виштовхує їх з клітки. Що вийшли катіони калію збільшують позитивний заряд на зовнішній поверхні клітинної мембрани. В результаті виходу іонів калію з клітки повинна була б незабаром зрівнятися їх концентрація всередині і поза клітиною. Однак цьому перешкоджає електрична сила відштовхування позитивних іонів калію від позитивно зарядженої зовнішньої поверхні мембрани.
Чим більше стає величина позитивного заряду на зовнішній поверхні мембрани, тим важче іонів калію проходити з цитоплазми через мембрану. Іони калію виходитимуть з клітки до тих пір, поки сила електричного відштовхування не стане рівною силі осмотичного тиску До +. При такому рівні потенціалу на мембрані вхід і вихід іонів калію з клітки знаходяться в рівновазі, тому електричний заряд на мембрані в цей момент називається калієвих рівноважним потенціалом. Для нейронів він дорівнює від -80 до -90 мВ.
Оскільки в спочиває клітці майже всі натрієві канали мембрани закриті, то іони Nа + надходять в клітину по концентраційному градієнту в незначній кількості. Вони лише в дуже малому ступені відшкодовують втрату позитивного заряду внутрішньої середовищем клітини, викликану виходом іонів калію, але не можуть цю втрату істотно компенсувати. Тому проникнення в клітину (витік) іонів натрію призводить лише до незначного зниження мембранного потенціалу, внаслідок чого мембранний потенціал спокою має декілька меншу величину в порівнянні з калієвих рівноважним потенціалом.
Таким чином, виходять з клітини катіони калію спільно з надлишком катіонів натрію в позаклітинній рідині створюють позитивний потенціал на зовнішній поверхні мембрани спочиває клітини.
У стані спокою плазматична мембрана клітини добре проникна для аніонів хлору. Аніони хлору, яких більше в позаклітинній рідині, дифундують всередину клітини і несуть з собою негативний заряд. Повного зрівнювання концентрацій іонів хлору зовні і всередині клітини не відбувається, тому що цьому перешкоджає сила електричного взаємного відштовхування однойменних зарядів. створюється хлорний рівноважний потенціал, при якому вхід іонів хлору в клітину і їх вихід з неї знаходяться в рівновазі.
Мембрана клітини практично непроникна для великих аніонів органічних кислот. Тому вони залишаються в цитоплазмі і спільно зі вступниками аніонами хлору забезпечують негативний потенціал на внутрішній поверхні мембрани спочиває нервової клітини.
Найважливіше значення мембранного потенціалу спокою полягає в тому, що він створює електричне поле, яке впливає на макромолекули мембрани і надає їх зарядженим групам певне положення в просторі. Особливо важливо те, що це електричне поле обумовлює закритий стан активаційних воріт натрієвих каналів і відкритий стан їх інактіваціонних воріт (рис. 61, А). Цим забезпечується стан спокою клітини і готовності її до порушення. Навіть відносно невелике зменшення мембранного потенціалу спокою відкриває активаційні «ворота» натрієвих каналів, що виводить клітку зі стану спокою і дає початок порушення.

Щоб провести сигнал від попередньої клітини до наступної, нейрон генерує електричні сигнали усередині себе. Твої рухи очима при читанні цього абзацу, відчуття м'якого крісла під попою, сприйняття музики з навушників і багато іншого засновані на тому, що всередині тебе проходять сотні мільярдів електричних сигналів. Такий сигнал може зародитися в спинному мозку і пройти до кінчика пальця ноги по довгому аксону. Або може подолати мізерно малу відстань в глибинах мозку, обмежуючись межами інтернейронов з короткими відростками. Будь-нейрон, який отримав сигнал, проганяє його через своє тіло і вирости, і цей сигнал має електричну природу.
Ще в 1859 році вчені змогли виміряти швидкість, з якою передаються ці електричні сигнали. Виявилося, що електрику, передане по живому аксону, принципово відрізняється від електричного струму в металах. По металевому проводу електричний сигнал передається зі швидкістю, близькою до швидкості світла (300 000 кілометрів на секунду), адже в металі багато вільних електронів. Однак, незважаючи на цю швидкість, сигнал відчутно слабшає, долаючи великі відстані. Якби по аксонах сигнали передавалися тим же способом, яким передаються в металах, то нервовий імпульс, що йде від нервового закінчення в шкірі великого пальця твоєї ноги, повністю загасав б, не досягаючи твого мозку - електричний опір органічної матерії занадто велике, а сигнал занадто слабкий .
Дослідження показали, що електрику передається по аксонах набагато повільніше, ніж по дротах, і що в основі цієї передачі лежить невідомий раніше механізм, в результаті якого сигнал поширюється зі швидкістю близько 30 метрів в секунду. Електричні сигнали, що йдуть по нервах, на відміну від сигналів, що йдуть по проводах, що не слабшають по ходу свого руху. Причина цього в тому, що нервові закінчення не пропускають через себе сигнал пасивно, просто дозволяючи наявними в них заряджених частинок передавати його один одному. Вони є в кожній своїй точці активним випромінювачем цього сигналу, ретранслюючи його, і докладний опис цього механізму потребують окремої глави. Таким чином, пожертвувавши високою швидкістю проведення нервових імпульсів, за рахунок активної передачі сигналу нейрон отримує гарантію того, що виник в великому пальці ноги сигнал досягне спинного мозку, анітрохи не втрачав сили.

Щоб спостерігати проходження електричної хвилі збудження, або потенціалу дії (action potential [ 'ækʃən pə'tenʃəl]), В живій клітині, досить простого пристрою: один кінець тонкого металевого дроту поміщається на зовнішню поверхню аксона сенсорного нейрона шкіри, а інший підводиться до самописця, чертящему лінію вгору при посиленні сигналу, і вниз - при ослабленні. Кожен дотик до шкіри викликає один або кілька потенціалів дії. При виникненні кожного потенціалу самописець малює вузький довгий пік.
Потенціал дії сенсорного нейрона триває всього лише близько 0,001 секунди і включає дві фази: швидкого наростання, що досягає піку, а потім майже настільки ж швидкого спаду збудження, що приводить до вихідного положення. І тут самописець повідомляє несподіваний факт: всі потенціали дії, що виникають в одній і тій же нервовій клітині, приблизно однакові. Це можна побачити на зображенні зліва: все піки, намальовані самописцем, мають приблизно одну і ту ж форму і амплітуду незалежно від того, наскільки сильним або тривалим був дотик до шкіри, їх викликала. Слабке погладжування або відчутний щипок будуть передані потенціалами дії однієї і тієї ж величини. Потенціал дії є постійний сигнал, який підпорядковується принципу «все або нічого»: після перевищення подразником якогось порогового значення виникає завжди приблизно однаковий сигнал, не більше і не менше, ніж звичайно. А якщо подразник менше порогового значення, то сигнал зовсім не буде передаватися: наприклад, можна так легко торкнутися шкіри кінчиком пера, що це дотик не відчува¹те.
Принцип «все або нічого» у виникненні потенціалу дії викликає нові питання. Як сенсорний нейрон повідомляє про силу подразника - сильне або слабке тиск, яскравий або тьмяне світло? Як він повідомляє про тривалість дії подразника? Нарешті, як нейрони відрізняють один тип сенсорної інформації від іншого - наприклад, як вони відрізняють дотик від болю, світла, запаху або звуку? І як вони відрізняють сенсорну інформацію для сприйняття від моторної інформації для дії?

Еволюція вирішила питання про те, як повідомити про силу подразника, за допомогою використання одного і того ж виду сигналів однієї і тієї ж величини: ця сила визначається частотою (Frequency [ 'friːkwənsɪ]), з якої випускаються потенціали дії. Слабкий подразник, наприклад легкий дотик до руки, призводить до випускання всього двох-трьох потенціалів дії в секунду, в той час як сильний тиск, як при щипки або ударі по ліктя, може викликати чергу з сотні потенціалів дії в секунду. При цьому тривалість відчуття визначається тривалістю виникнення потенціалів дії.
Чи використовують нейрони різні електричні коди, повідомляючи мозку, що несуть інформацію про різні подразники, таких як біль, світло або звук? Виявилося, що ні! Це дивно, але між потенціалами дії, що генеруються нейронами з різних сенсорних систем (наприклад, зорової або тактильної), різниця досить незначна! Таким чином, характер і природа відчуття не залежать від відмінностей в потенціалах дії (що відкриває досить захоплюючу перспективу для роздумів на тему «матриці» з однойменного фільму). Нейрон, що передає слухову інформацію, влаштований точно так же, як нейрон з зорової нервової ланцюга, і проводять вони одні й ті ж потенціали дії, одним і тим же способом. Без знання, до якої нервової ланцюга належить конкретний нейрон, тільки з аналізу його функціонування неможливо визначити, яку інформацію він несе.
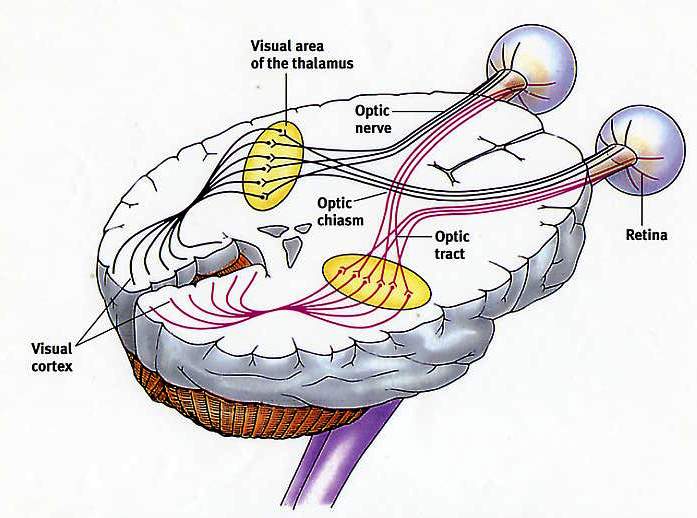
Природа переданої інформації залежить насамперед від типу порушуваних нервових волокон і специфічних систем мозку, з якими ці волокна пов'язані. Відчуття кожного типу передаються за своїми провідних шляхах, і різновид переданої нейроном інформації залежить саме від шляху, до складу якого входить цей нейрон. У будь-якому сенсорному провідному шляху інформація передається від першого сенсорного нейрона (рецептора, що реагує на зовнішній подразник, наприклад дотик, запах або світло) до спеціалізованих нейронам в спинному або головному мозку. Таким чином, зорова інформація відрізняється від слуховий лише тим, що передається по іншим провідним шляхам, що починається в сітківці ока і закінчується в ділянці мозку, який відповідає за візуальне сприйняття.
Сигнали, що посилаються від моторних нейронів мозку до м'язів, також майже ідентичні переданим по сенсорним нейронам від шкіри в мозок. Вони підпорядковуються тим же принципом «все або нічого», так само передають інтенсивність сигналу за допомогою частоти потенціалів дії, і так же результат сигналу залежить тільки від того, в яку нервову ланцюг включений цей нейрон. Таким чином, швидка низка потенціалів дії, що йде за певним провідного шляху, викликає саме рух твоїх пальців, а не, скажімо, сприйняття різнокольорових вогнів, лише тому, що даний шлях пов'язаний з м'язами рук, а не з сітківкою очей.

Універсальність потенціалів дії не обмежується схожістю їх прояви в різних нейронах, що знаходяться в межах одного організму. Вони настільки однакові у різних тварин, що навіть навчений досвідом дослідник не здатний точно відрізнити запис потенціалу дії нервового волокна кита, миші, мавпи або його наукового керівника. Проте потенціали дії в різних клітинах не є ідентичними: невелика різниця в їх амплітуді і тривалості все ж є, і твердження «все потенціали дії однакові» так само неточно, як і «все бугенвиллии однакові».
Отже, кожен нейрон передає сигнал через своє тіло і відростки одним і тим же чином. Все розмаїття інформації, одержуваної нами від сенсорних нейронів, всі рухи, які може здійснювати наше тіло - результат передачі єдиного типу сигналів усередині нейронів. Залишилася «дрібниця»: зрозуміти, що ж це за сигнал і як він передається.

Ми звично відокремлюємо все, що вважаємо живою природою, в тому числі і себе самих, від «неживих» речей, в тому числі металів і передається через них електричного струму. Тим дивніше усвідомлювати, що в наших тілах метали не просто присутні - вони необхідні, без них тіло не зможе існувати. Електричний струм - явище не разовий, а безперервно виникає в сотні мільярдів нейронів, пронизали своїми відростками все наше тіло. Прямо зараз ти можеш відчути найрізноманітніші ознаки його присутності: то, що ти усвідомлюєш цей текст, є результат незліченних передач електричного струму. Почуття голоду і задоволення від запаху готується їжі, саме сприйняття цього запаху, дотик залетів у вікно вітру до твоїй шкірі ... Перераховувати можна нескінченно. І бажання зрозуміти, яким же чином все це відбувається, також складається з виникаючих в нейронах електричних імпульсів.
Так як метою цього розділу є повідомлення лише самої загальної інформації про проходження нервового імпульсу, то тут же необхідно розглянути те середовище, в якій він виникає, ті умови в клітці, які роблять можливим його виникнення і передачу. Тому варто почати з вивчення плацдарму, на якому будуть розвиватися події, а саме з нейрона в стані спокою (dormant state [ 'dɔːmənt steɪt]).

Ще в середині минулого століття вчені знайшли спосіб встановити, в якій частині нейрона існує електричний заряд. Для цього використовують вольтметр (voltmeter [ 'vəultˌmiːtə]) (Прилад для вимірювання напруги електричного поля) З двома електродами. Один електрод поміщають всередину нейрона, розташовуючи його близько до клітинної мембрани, а другий електрод знаходиться в навколишньому нейрон середовищі, з іншого боку тієї ж мембрани. Вольтметр показує, що з різних сторін клітинної мембрани існують електричні заряди, Негативний всередині клітини і позитивний зовні. Існування таких різнополюсних електричних зарядів по обидві сторони мембрани створює електричне поле, важливою характеристикою якого є потенціал. Потенціал, кажучи простою мовою, це здатність виконувати роботу, наприклад роботу з перенесення зарядженої частинки з місця на місце. Чим більше негативних зарядів накопичилося по одну сторону, і чим більше позитивних - по іншу сторону мембрани, тим сильніше створюване ними електричне поле, і тим з більшою силою вони здатні перетягувати туди-сюди заряджені частинки. Різницю між зовнішнім і внутрішнім електричними зарядами називають мембранним потенціалом (membrane potential [ 'membreɪn pə'tenʃəl]) Спокою. Для нейрона він дорівнює приблизно 70 мВ (мілівольт), тобто 70 тисячних вольта або сім сотих вольта. Для порівняння, різниця потенціалів в батарейці АА дорівнює 1,5 вольта - в 20 разів більше. Тобто мембранний потенціал спокою нейрона всього лише в 20 разів слабкіше, ніж між клемами батарейки АА - досить великий, виходить. Електричний потенціал існує тільки на мембрані, і в інших своїх частинах нейрон електрично нейтральний.
Якщо написати більш точно, то мембранний потенціал спокою нейрона дорівнює -70 мВ (мінус сімдесят мілівольт). Знак мінус означає лише те, що негативний заряд знаходиться саме всередині клітини, а не зовні, і таким чином створюється електричне поле здатне перетягувати через мембрану всередину клітини позитивно заряджені іони.
Дійові особи в створенні мембранного потенціалу спокою:

1 . В клітинній мембрані нейрона існують канали, за якими несуть електричний заряд іони можуть подорожувати крізь неї. При цьому мембрана не є всього лише пасивної «перегородкою» між внутрішнім середовищем нейрона і навколишнього його міжклітинної рідиною: спеціальні білки, впроваджені в плоть мембрани, відкривають і закривають ці канали, і таким чином мембрана контролює проходження йонів - атомів, що мають електричний заряд. Накопичуючи негативно заряджені іони всередині клітини, нейрон збільшує кількість негативних зарядів всередині, тим самим приводячи до збільшення позитивних зарядів зовні, і таким чином посилюється електричний потенціал. Так як протон має позитивний заряд, а електрон негативний, то при надлишку протонів виходить позитивно заряджений іон, а при надлишку електронів - негативно заряджений. Якщо хочеться більш докладної інформації про атомах і іони, можна повернутися в. Важливо розуміти, що мембранний потенціал існує саме на кордоні клітинної мембрани, а рідини в цілому всередині і поза нейрона залишаються електрично нейтральними. Іони, для яких мембрана проникна, залишаються поблизу неї, оскільки позитивні і негативні заряди взаємно притягуються один до одного. В результаті зовні мембрани утворюється шар «сидять» на ній позитивних іонів, а всередині - негативних. Таким чином, мембрана відіграє роль електричної ємності, що розділяє заряди, всередині якої є електричне поле. Мембрана, тому, є природним конденсатором.
2 . негативно заряджені протеїни, Що знаходяться всередині нейрона біля внутрішньої поверхні мембрани. Заряд протеїнів завжди залишається одним і тим же і є тільки частиною загального заряду внутрішньої поверхні мембрани. На відміну від іонів, протеїни не можуть виходити з клітини і заходити в неї - для цього вони занадто великі. Загальний заряд змінюється в залежності від кількості знаходяться біля мембрани позитивно заряджених іонів, концентрація яких може змінюватися за рахунок їх переходу з клітки назовні, і ззовні всередину.
3 . позитивно заряджені іони калію (К +) можуть вільно переміщатися між внутрішнім і зовнішнім середовищем, коли нейрон знаходиться в стані спокою. Переміщаються вони через постійно відкриті проточні калієві канали (flow potassium passage), Через які можуть пройти тільки іони К +, і ніщо інше. Проточними називаються канали, які не мають воріт, а значить відкриті при будь-якому стані нейрона. Усередині клітини іонів калію набагато більше, ніж зовні. Це відбувається за рахунок постійної роботи натрій-калієвого насоса (про нього буде розказано нижче), тому в стані спокою нейрона іони К + починають переміщатися в зовнішнє середовище, оскільки концентрація одного і того ж речовини прагне вирівнятися в загальній системі. Якщо ми в басейн з водою в одному кутку виллємо якусь речовину, то його концентрація в цьому кутку буде дуже великий, а в інших частинах басейну - нульовий або дуже маленькою. Однак, через деякий час ми виявимо, що концентрація цієї речовини вирівнялася по всьому басейну за рахунок броунівського руху. У цьому випадку говорять про «парціальному тиску» того чи іншого речовини, будь це рідина або газ. Якщо в одному кутку басейну буде вилитий спирт, то утворюється велика різниця в концентрації спирту між цим кутом і іншим басейном. Виникне парціальний тиск молекул спирту, і вони поступово розподіляться рівномірно по басейну так, що парціальний тиск зникне, оскільки концентрація молекул спирту всюди вирівняється. Таким чином, іони К + забирають з собою позитивний заряд з нейрона, йдучи назовні за рахунок парціального тиску, яке сильніше, ніж сила тяжіння негативно заряджених протеїнів, в тому випадку, якщо різниця в концентрації іонів всередині і зовні клітини досить велика. Так як всередині залишаються негативно заряджені протеїни, то таким чином на внутрішній стороні мембрани формується негативний заряд. Для ясного розуміння роботи клітинних механізмів важливо пам'ятати, що незважаючи на постійне витікання іонів калію з клітки, всередині нейрона їх завжди більше, ніж зовні.
4 . позитивно заряджені іони натрію (Na +) знаходяться із зовнішнього боку мембрани і створюють там позитивний заряд. Під час фази спокою нейрона натрієві канали клітини закриті, І Na + не можуть пройти всередину, а їх концентрація зовні підвищується за рахунок роботи натрій-калієвого насоса, що виводить їх з нейрона.
5 . роль негативно заряджених іонів хлору (Cl -) і позитивно заряджених іонів кальцію (Ca 2+) для створення мембранного потенціалу невелика, тому їх поведінка поки залишиться за кадром.
Формування мембранного потенціалу спокою проходить в два етапи:
етап I. створюється невелика (-10 мВ) різниця потенціалів за допомогою натрій-калієвого насоса.

На відміну від інших каналів мембрани, натрій-калієвий канал здатний пропускати через себе і іони натрію, і іони калію. Причому Na + може пройти крізь нього тільки з клітини назовні, а К + зовні всередину. Один цикл роботи цього каналу включається в себе 4 етапи:
1 . «Ворота» натрій-калієвого каналу відкриті тільки з внутрішньої сторони мембрани, і туди заходять 3 Na +

2 . присутність Na + всередині каналу впливає на нього так, що він може частково зруйнувати одну молекулу АТФ (ATP) ( аденозинтрифосфату), (adenosine triphosphate [ə'dɛnəsiːn trai'fɔsfeɪt]) Що є «акумулятором» клітини, запасаються енергію і віддають її при необхідності. При такому частковому руйнуванні, що полягає в відщепленні від кінця молекули однієї фосфатної групи PO 4 3-, виділяється енергія, яка як раз і витрачається на перенесення Na + у зовнішній простір.
3 . коли канал відкривається для того, щоб Na + вийшов назовні, він залишається відкритим, і в нього потрапляють два іони К + - їх притягують негативні заряди протеїнів зсередини. Те, що в каналі, що вміщає три іона натрію, поміщається всього два іона калію, цілком логічно: атом калію має більший діаметр.
4 . присутність іонів калію тепер в свою чергу впливає на канал так, що зовнішні «ворота» закриваються, а внутрішні відкриваються, і К + надходять у внутрішнє середовище нейрона.
Таким чином працює натрій-калієвий насос, «обмінюючи» три іона натрію на два іона калію. Так як електричний заряд у Na + і К + однаковий, виходить що з клітки виводиться три позитивних заряду, а всередину потрапляє тільки два. За рахунок цього внутрішній позитивний заряд клітинної мембрани знижується, а зовнішній - збільшується. До того ж створюється різниця в концентрації Na + і К + по різні боки мембрани:
\u003d) Зовні клітини виявляється багато іонів натрію, а всередині - мало. При цьому натрієві канали закриті, і потрапити назад в клітку Na + не може, і далеко від мембрани він не йде, так як притягається існуючим з внутрішньої сторони мембрани негативним зарядом.
\u003d) Всередині клітини багато іонів калію, а от зовні їх мало, і це призводить до витікання До + з клітки через відкриті під час фази спокою нейрона калієві канали.

етап II формування мембранного потенціалу спокою якраз заснований на цьому витіканні іонів калію з нейрона. На малюнку ліворуч показано іонний склад мембрани на початку другого етапу формування потенціалу спокою: безліч До + і негативно заряджених протеїнів (позначених А 4) всередині, і обліпили мембрану зовні Na +. Переміщаючись в зовнішнє середовище, іони калію забирають з клітки свої позитивні заряди, при цьому сумарний заряд внутрішньої мембрани знижується. Так само як позитивні іони натрію, що витекла з клітки іони калію залишаються зовні мембрани, притягує внутрішнім негативним зарядом, і зовнішній позитивний заряд мембрани складається з суми зарядів Na + і К +. Незважаючи на витікання через проточні канали, усередині клітини іонів калію завжди більше, ніж зовні.
Виникає питання: чому іони калію НЕ продовжують витікати назовні до того моменту, поки їх кількість всередині клітини і поза нею не стане однаковим, тобто до тих пір, поки не зникне парціальний тиск, що створюється цими іонами? Причина цього полягає в тому, що коли К + залишають клітку, зовні збільшується позитивний заряд, а всередині утворюється надлишок негативного заряду. Це знижує бажання іонів калію виходити з клітки, адже зовнішній позитивний заряд їх відштовхує, а внутрішній негативний притягує. Тому через якийсь час До + перестають витікати незважаючи на те, що у зовнішньому середовищі їх концентрація нижче, ніж у внутрішній: вплив зарядів по різні боки мембрани перевищує силу парціального тиску, тобто перевищує прагнення К + розподілитися рівномірно в рідини всередині і поза нейрона. У момент досягнення цієї рівноваги мембранний потенціал нейрона і зупиняється приблизно на -70 мВ.
Як тільки нейроном досягнутий мембранний потенціал спокою, він готовий для виникнення і проведення потенціалу дії, про який мова піде в наступній цитологічної чолі.
Таким чином, підсумуємо: Нерівномірність розподілу іонів калію і натрію по обидві сторони мембрани викликана дією двох конкуруючих сил: а) силою електричного тяжіння і відштовхування, і б) силою парціального тиску, що виникає при різниці в концентраціях. Робота цих двох конкуруючих сил протікає в умовах існування по-різному влаштованих натрієвих, калієвих і натрієво-калієвого каналів, які виступають в ролі регуляторів дії цих сил. Калійний канал є проточним, тобто він завжди відкритий в стані спокою нейрона, так що іони К + можуть спокійно ходити туди-сюди під впливом сил електричного відштовхування / тяжіння і під впливом сили, викликані парціальним тиском, тобто різницею в концентрації цих іонів. Натрієвий канал завжди закритий в стані спокою нейрона, так що через них іони Na \u200b\u200b+ ходити не можуть. І, нарешті, натрієво-калієвий канал, влаштований так, що він працює як насос, який при кожному циклі виганяє три іона натрію назовні, і заганяє два іона калію всередину.
Вся ця конструкція і забезпечує виникнення мембранного потенціалу спокою нейрона: тобто стану, при якому досягається дві речі:
а) всередині є негативний заряд, а зовні - позитивний.
б) всередині багато іонів К +, що обліпили негативно заряджені частини протеїнів, і таким чином виникає калиевое парціальний тиск - прагнення іонів калію вийти назовні для вирівнювання концентрації.
в) зовні багато іонів Na +, що утворюють частково пари з іонами Cl -. І таким чином виникає натриевое парціальний тиск - прагнення іонів натрію увійти всередину клітини для вирівнювання концентрації.
В результаті роботи калієво-натрієвого насоса ми отримуємо три сили, що існують на мембрані: силу електричного поля і силу двох парціальних тисків. Ці сили і починають працювати, коли нейрон виходить зі стану спокою.
Історія відкриття
У 1902 році Юліус Бернштейн висунув гіпотезу, згідно з якою клітинна мембрана пропускає всередину клітини іони К +, і вони накопичуються в цитоплазмі. Розрахунок величини потенціалу спокою за рівнянням Нернста для калієвого електрода задовільно збігся з виміряним потенціалом між саркоплазмою м'язи і навколишнім середовищем, Який становив близько - 70 мВ.
Відповідно до теорії Ю. Бернштейна, при порушенні клітини її мембрана ушкоджується, і іони К + випливають з клітки по концентраційному градієнту до тих пір, поки потенціал мембрани не стає рівним нулю. Потім мембрана відновлює свою цілісність, і потенціал повертається до рівня потенціалу спокою. Це твердження, що відноситься скоріше до потенціалу дії, було спростовано Ходжкином і Хакслі в 1939 році.
Теорію Бернштейна щодо потенціалу спокою підтвердив Кеннет Стюарт Коул (Kenneth Stewart Cole), іноді його ініціали помилково пишуть як K.C. Cole, через його прізвиська, Кейсі ( «Kacy»). ПП і ПД зображені на відомій ілюстрації Коула і Curtis, 1939. Цей малюнок став емблемою Membrane Biophysics Group of the Biophysical Society (див. Ілюстрацію).
загальні положення
Для того, щоб на мембрані підтримувалася різниця потенціалів, необхідно, щоб була певна різниця концентрації різних іонів всередині і зовні клітини.
Концентрації іонів в клітці скелетного м'яза і в позаклітинному середовищі
Потенціал спокою для більшості нейронів становить величину порядку -60 мВ - -70 мВ. У клітин незбудливих тканин на мембрані також є різниця потенціалів, різна для клітин різних тканин і організмів.
Формування потенціалу спокою
ПП формується в два етапи.
Перший етап: створення незначною (-10 мВ) отріцательності всередині клітини за рахунок нерівного асиметричного обміну Na + на K + в співвідношенні 3: 2. У результаті цього клітку залишає більше позитивних зарядів з натрієм, ніж повертається в неї з калієм. Така особливість роботи натрій-калієвого насоса, який здійснює взаємообмін цих іонів через мембрану з витратами енергії АТФ, забезпечує його Електрогене.
Результати діяльності мембранних іонних насосів-обмінників на першому етапі формування ПП такі:
1. Дефіцит іонів натрію (Na +) в клітці.
2. Надлишок іонів калію (K +) в клітці.
3. Поява на мембрані слабкого електричного потенціалу (-10 мВ).
Другий етап: створення значної (-60 мВ) отріцательності всередині клітини за рахунок витоку з неї через мембрану іонів K +. Іони калію K + залишають клітку і забирають з собою з неї позитивні заряди, доводячи негативність до -70 мВ.
Отже, мембранний потенціал спокою - це дефіцит позитивних електричних зарядів всередині клітини, що виникає за рахунок витоку з неї позитивних іонів калію і Електрогене дії натрій-калієвого насоса.
Див. також
Примітки
посилання
Дуділи Й., Рюегг Й., Шмідт Р. і ін. Фізіологія людини: в 3-х томах. Пер. з англ / під ред Р. Шмідта і Г. Тевса. - 3. - М.: Світ, 2007. - Т. 1. - 323 с илл. с. - 1500 екз. - ISBN 5-03-000575-3
Wikimedia Foundation. 2010 року.
Дивитися що таке "Потенціал спокою" в інших словниках:
ПОТЕНЦІАЛ СПОКОЮ, електричний потенціал між внутрішньою і зовнішньою середовищем клітини, що виникає на її мембрані; у нейронів і м'язових клітин досягає величини 0,05 0,09 В; виникає через нерівномірний розподіл і накопичення іонів по різні ... енциклопедичний словник
Мембранний потенціал спокою, різниця потенціалів, яка існує у живих клітин в стані физиол. спокою, між їх цитоплазмою і позаклітинної рідиною. У нервових і м'язових клітин П. п. Варіює зазвичай в діапазоні 60 90 мВ, причому внутр. сторона ...
потенціал спокою - напруга спокою - [Я.Н.Лугінскій, М.С.Фезі Жилінський, Ю.С.Кабіров. Англо російський словник з електротехніки та електроенергетиці, Москва, 1999 г.] Тематики електротехніка, основні поняття Синоніми напруга спокою EN rest potentialresting ... ... Довідник технічного перекладача
потенціал спокою - Rest (ing) Potential Потенціал спокою Потенціал, який існує між середовищем, в якій знаходиться клітина, і її вмістом ... Тлумачний англійсько-російський словник по нанотехнології. - М.
потенціал спокою - Потенціал неактивного нейрона. Називається також мембранним потенціалом ... Психологія відчуттів: глосарій
потенціал спокою - різниця потенціалу між вмістом клітини і позаклітинної рідиною. У нервових клітинах п.п. бере участь в підтримці готовності клітини до порушення. * * * Мембранний біоелектричний потенціал (близько 70мВ) в нервовій клітині, що знаходиться в ... ... Енциклопедичний словник з психології та педагогіки
потенціал спокою - - різниця електричного заряду між зовнішньою і внутрішньою поверхнями мембрани в стані фізіологічного спокою клітини, реєстрований до початку дії подразника ... Словник термінів по фізіології сільськогосподарських тварин
Мембранний потенціал, який реєструється до початку дії подразника ... Великий медичний словник
- (фізіологічний) різниця потенціалів між вмістом клітини (волокна) і позаклітинної рідиною; стрибок потенціалу локалізується на поверхневій мембрані, при цьому її, внутрішня сторона заряджена електронегативно по відношенню до ... ... Велика Радянська Енциклопедія
Швидке коливання (спайк) мембранного потенціалу, що виникає при порушенні нервових, м'язових, недо яких залізистих і ростить, клітин; елект. сигнал, що забезпечує швидку передачу інформації в організмі. Підкоряється правилу «все або нічого» ... ... Біологічний енциклопедичний словник
книги
- 100 способів змінити життя. Частина 1, Парфентьева Лариса. Про книгу Збірник надихаючих історій про те, як змінити життя на краще, від людини, яка зуміла розгорнути своє власне життя на 180 градусів. Ця книга народилася з щотижневої рубрики ...
Встановлено, що найбільш важливими іонами, що визначають мембранні потенціали клітин, є неорганічні іони К +, Na +, СГ, а також в ряді випадків Са 2 +. Добре відомо, що концентрації цих іонів в цитоплазмі і в міжклітинної рідини розрізняються в десятки разів.
З табл. 11.1 видно, що концентрація іонів К + всередині клітини в 40-60 разів більше, ніж в міжклітинної рідини, тоді як для Na + і СГ розподіл концентрацій протилежне. Нерівномірний розподіл концентрацій цих іонів по обидві сторони мембрани забезпечується як їх різною проникністю, так і сильним електричним полем мембрани, яке визначається її потенціалом спокою.
Дійсно, в стані спокою сумарний потік іонів через мембрану дорівнює нулю, і тоді з рівняння Не- рнста - Планка слід, що
Таким чином, в спокої градієнти концентрації - і
електричного потенціалу - на мембрані спрямовані
протилежно один одному і тому в спочиває клітці висока і постійна різниця концентрацій основних іонів забезпечує підтримку на мембрані клітини електричної напруги, яке і називають рівноважним мембранним потенціалом.
У свою чергу виникає на мембрані потенціал спокою перешкоджає виходу іонів з клітки К + і надмірного входу в неї СГ, підтримуючи тим самим їх концентраційні градієнти на мембрані.
Повний вираз для мембранного потенціалу, що враховує потоки дифузії цих трьох видів іонів, було отримано Гольдманом, Ходжкіна і Катц:
де Р до, P Na, Р С1 - проникність мембрани для відповідних іонів.
Рівняння (11.3) з високою точністю визначає мембранні потенціали спокою різних клітин. З нього випливає, що для мембранного потенціалу спокою важливі не абсолютні величини проницаемостей мембрани для різних іонів, а їх відносини, так як, розділивши обидві частини дробу під знаком логарифма, наприклад, на Р до, ми перейдемо до відносних проникність іонів.
У тих випадках, коли проникність одного з цих іонів значно більше, ніж інших, рівняння (11.3) переходить в рівняння Нернста (11.1) для цього іона.
З табл. 11.1 видно, що мембранний потенціал спокою клітин близький до потенціалу Нернста для іонів К + і СВ, але значно відрізняється від нього по Na +. це свідчить
0 тому, що в спокої мембрана добре проникна для іонів К + і СГ, тоді як для іонів Na + її проникність дуже низька.
Незважаючи на те що рівноважний потенціал Нернста для СГ найбільш близький до потенціалу спокою клітини, останній має переважно калиевую природу. Це обумовлено тим, що висока внутрішньоклітинна концентрація К + не може істотно зменшитися, так як іони К + повинні врівноважувати всередині клітини об'ємний негативний заряд аніонів. Внутрішньоклітинні аніони являють собою в основному великі органічні молекули (Білки, залишки органічних кислот іт.п.), які не можуть пройти через канали в клітинній мембрані. Концентрація цих аніонів в клітці практично постійна і їх сумарний негативний заряд перешкоджає значного виходу калію з клітки, підтримуючи разом з Na-K-насосом його високу внутрішньоклітинну концентрацію. Однак основна роль в первісному встановленні всередині клітини високої концентрації іонів калію і низької концентрації іонів натрію належить Na-K-насоса.
Розподіл іонів С1 встановлюється відповідно до мембранним потенціалом, оскільки в клітці немає спеціальних механізмів підтримки концентрації СГ. Тому внаслідок негативного заряду хлору його розподіл виявляється зворотним по відношенню до розподілу калію на мембрані (див. Табл. 11.1). Таким чином, концентраційні дифузії К + з клітини і С1 в клітку практично врівноважуються мембранним потенціалом спокою клітини.
Що стосується Na +, то в спокої його дифузія направлена \u200b\u200bв клітку під дією як градієнта концентрації, так і електричного поля мембрани і вхід Na + в клітину обмежується в спокої тільки малої проникністю мембрани для натрію (закриті натрієві канали). Дійсно, Ходжкин і Катц експериментально встановили, що в стані спокою проникності мембрани аксона кальмара для К +, Na + і СГ відносяться як 1: 0,04: 0,45. Таким чином, в стані спокою клітинна мембрана малопроніцаема тільки для Na +, а для СГ вона проникна майже так само добре, як і для К +. У нервових клітинах проникність для СГ зазвичай нижче, ніж для К +, але в м'язових волокнах проникність для СГ навіть дещо переважає.
Незважаючи на малу проникність клітинної мембрани для Na + в спокої, існує, хоча і вельми малий, пасивний перенесення Na + в клітину. Цей струм Na + мав би приводити до зниження різниці потенціалів на мембрані і до виходу К + з клітини, а це призводило б в кінцевому підсумку до вирівнювання концентрацій Na + і К + по обидві сторони мембрани. Цього не відбувається завдяки роботі Na + - К + -насоса, компенсуючого струми витоку Na + і К + і підтримує таким чином нормальні значення внутрішньоклітинних концентрацій цих іонів і, отже, нормальну величину потенціалу спокою клітини.
Для більшості клітин мембранний потенціал спокою становить (-бО) - (- ЮО) мВ. На перший погляд може здатися, що це мала величина, але треба врахувати, що товщина мембрани теж мала (8-10 нм), так що напруженість електричного поля в клітинній мембрані величезна і становить близько 10 млн вольт на 1 м (або 100 кВ на 1 см):
Повітря, наприклад, не витримує такої напруженості електричного поля (електричний пробій в повітрі настає при 30 кВ / см), а мембрана витримує. Це нормальна умова її діяльності, оскільки саме таке електричне поле необхідно для підтримки різниці концентрацій іонів натрію, калію і хлору на мембрані.
Величина потенціалу спокою, різна у клітин, може змінюватися при зміні умов їх життєдіяльності. Так, порушення біоенергетичних процесів в клітині, що супроводжується падінням внутрішньоклітинного рівня макро- ергічних сполук (зокрема, АТФ), перш за все виключає компоненту потенціалу спокою, пов'язану з роботою Ма + -К + -АТФ-ази.
Пошкодження клітини призводить зазвичай до підвищення проникності клітинних мембран, в результаті чого відмінності в проникності мембрани для іонів калію і натрію зменшуються; потенціал спокою при цьому зменшується, що може викликати порушення ряду функцій клітини, наприклад збудливості.
- Оскільки внутрішньоклітинна концентрація калію підтримується майже постійною, то навіть відносно невеликі зміни позаклітинної концентрації К * можуть помітно впливати на потенціалпокоя і на діяльність клітини. Подібні зміни концентрації К "в плазмі крові відбуваються при деяких патологіях (наприклад, пріпочечной недостатності).